2024年广东高考生物学真题第18、19、21题的文献溯源及其分析
01
课件目录
点击课件即可预览课件详情
必修2 遗传与进化
第1章 遗传因子的发现
选择性必修1 稳态与调节
第1章 人体的内环境与稳态
第1节 细胞生活的环境
第2节 内环境的稳态
章末质量检测卷1
第2章 神经调节
第1节 神经调节的结构基础
第2节 神经调节的基本方式
第3节 神经冲动的产生和传导
第4节 神经系统的分级调节
第5节 人脑的高级功能
章末质量检测卷2
第3章 体液调节
第1节 激素与内分泌系统
第2节 激素调节的过程
第3节 体液调节与神经调节的关系
章末质量检测卷3
第4章 免疫调节
第1节 免疫系统的组成和功能
第2节 特异性免疫
第3节 免疫失调
第4节 免疫学的应用
章末质量检测卷4
第5章 植物生命活动的调节
第1节 植物生长素
第2节 其他植物激素
第3节 植物生长调节剂的应用
第4节 环境因素参与调节植物的生命活动
章末质量检测卷4
全册质量检测卷3
选择性必修2 生物与环境
第1章 种群及其动态
第1节 种群的数量特征
第2节 种群数量的变化
第3节 影响种群数量变化的因素
章末质量检测卷1
第2章 群落及其演替
第1节 群落的结构
第2节 群落的主要类型
第3节 群落的演替
章末质量检测卷2
第3章 生态系统及其稳定性
第1节 生态系统的结构
第2节 生态系统的能量流动
第3节 生态系统的物质循环
第4节 生态系统的信息传递
第5节 生态系统的稳定性
章末质量检测卷3
第4章 人与环境
第1节 人类活动对生态环境的影响
第2节 生物多样性及其保护
第3节 生态工程
章末质量检测卷4
全册质量检测卷4
选择性必修3 生物技术与工程
第1章 发酵工程
第1节 传统发酵技术的应用
第2节 微生物的培养技术及应用(第一课时)
第2节 微生物的培养技术及应用(第二课时)
第3节 发酵工程及其应用
章末质量检测卷1
第2章 细胞工程
第1节 植物细胞工程
第2节 动物细胞工程(第1课时 动物细胞培养)
第2节 动物细胞工程(第2课时 动物细胞融合技术与单克隆抗体)
第2节 动物细胞工程(第3课时 动物体细胞核移植技术和克隆动物)
第3节 胚胎工程(第1课时 胚胎工程的理论基础)
第3节 胚胎工程(第2课时 胚胎工程技术及其应用)
章末质量检测卷2
第3章 基因工程
第1节 重组DNA技术的基本工具
第2节 基因工程的基本操作程序(第1课时)
第2节 基因工程的基本操作程序(第2课时)
第2节 基因工程的基本操作程序(第3课时)
第3节 基因工程的应用
第4节 蛋白质工程的原理和应用
章末质量检测卷3
第4章 生物技术的安全性与伦理问题
第1节 转基因产品的安全性
第2节 关注生殖性克隆人
第3节 禁止生物武器
章末质量检测卷4
全册质量检测卷5
02
课件获取
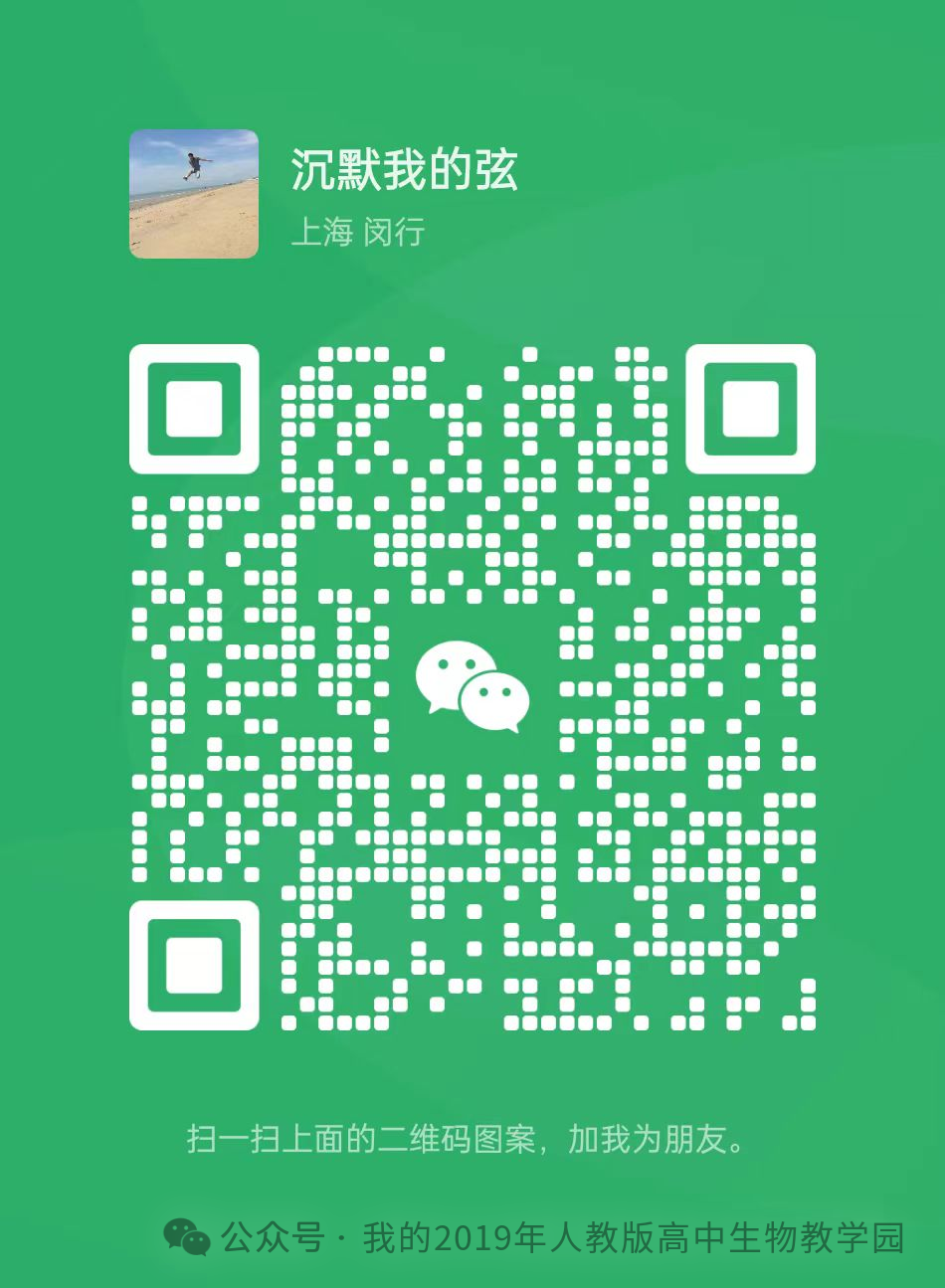
原题再现;
21.(13分)驹形杆菌可合成细菌纤维素(BC)并将其分泌到胞外组装成膜。作为一种性能优异的生物材料,BC膜应用广泛。研究者设计了酪氨酸酶(可催化酪氨酸形成黑色素)的光控表达载体,将其转入驹形杆菌后构建出一株能合成BC膜并可实现光控染色的工程菌株,为新型纺织原料的绿色制造及印染工艺升级提供了新思路(图一)|。
回答下列问题:
(1)研究者优化了培养基的(答两点)等营养条件,并控制环境条件,大规模培养工程菌株后可在气液界面处获得BC菌膜(菌体和BC膜的复合物)。
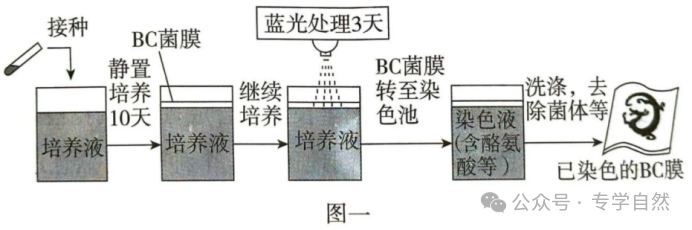
(2)研究者利用T7噬菌体来源的RNA聚合酶(T7RNAP)及蓝光光敏蛋白标签,构建了一种可被蓝光调控的基因表达载体(光控原理见图二a,载体的部分结构见图二b)。构建载体时,选用了通用型启动子PBAD(被工程菌RNA聚合酶识别)和特异型启动子PT7(仅被T7RNAP识别)。为实现蓝光控制染色,启动子①②及③依次为。理由是。
(3)光控表达载体携带大观霉素(抗生素)抗性基因。长时间培养时在培养液中加入大观霉素,其作用为(答两点)。
(4)根据预设的图案用蓝光照射已长出的BC菌膜并继续培养一段时间,随后将其转至染色池处理,发现只有经蓝光照射的区域被染成黑色,其原因是。
(5)有企业希望生产其他颜色图案的BC膜。按照上述菌株的构建模式提出一个简单思路:。
参考答案(1)碳源、氮源
(2)P17、PBAD和PBAD基因2和3正常表达无活性产物,被蓝光激活后再启动基因1表达
(3)抑制杂菌,去除丢失质粒的菌株
(4)该处细胞中T7RNAP激活,酪氨酸酶表达并合成黑色素
(5)将酪氨酸酶替换成催化其他色素合成的酶(或将酪氨酸酶替换成不同颜色蛋白)
情境分析:如果有老师曾经参加过或者带学生参加过iGEM比赛,这道题会不会觉得似曾相识?没错,命题情境来自合成生物学,是重组DNA技术发展到今天比较前沿的应用领域之一。如果说蛋白质工程中改造基因获得新的蛋白质,属于第二代基因工程。那么合成生物学中通过模块化设计基因线路来生产所需产品,就属于第三代基因工程。放一张某版本教科书对这一领域做的介绍:
下面分析一下设问:
第(1)问设问考察微生物培养中的营养条件,本意并不是记忆层面的考察,解题时应首先抓住关键信息——目的是获得菌体和纤维素膜,因此从元素组成的角度考虑,需要优化的是碳源、氮源。然而考生实际作答时,可能只需要从脑海中提取教材相关内容,不做过多分析也能拿分。
第(2)问是考察如何基于给定的光遗传学元件来设计基因线路,最终使工程菌产生黑色素,这一问思维含金量高,也是难度最大的一问。实际解题时,可通过枚举可能的启动子组合,并在草稿纸上逐一演绎推理后排除不符合要求的组合。本问的难度主要在于情境新颖读不懂题(大多数同学高三阶段可能做了1年的练习都不曾碰到过1道,哪怕有半点相似的题目吧?)、字数繁多、加上所剩时间不多(可能只剩10分钟,前面大题还有空的),高度紧张的情况下,能做对这一问的注定只是少数人。
第(3)问考察基因工程中标记基因的作用,较为基础。注意流传版本的答案给的动词描述是“去除”,考虑的应该是工程菌繁殖过程中质粒复制不同步可能出现丢失的问题。
第(4)问考察如何基于设计好的基因线路,生产想要的图案。放一张原文的图,只有蓝光照射的区域才被染成了黑色:
第(5)问考察的是这一工程技术的应用价值,可以说是一种展望,源自原文的Discussion部分。
总的来说,笔者认为21题是24年广东卷5道大题中质量最高的题目,情境完全改编自真实、高质量的科学研究,且非常新颖。知识点融合了基因工程与发酵工程,设问层层递进,思维层级涵盖了理解、应用、分析、创造,4个学科核心素养在设问中都有落实。美中不足的是(2)(4)小问考察的内容略有重复,笔者在解第(2)问时,第(4)问的答案已经出来了。此外,难度比较大的设问出现在前边,无形中增加了学生的解题负担。许多考生做到21题可能只剩10分钟,看到第(2)问看不懂,后边的问题即使简单可能也直接放弃或乱填了。
教学启示许多老师在复习基因工程的时候,可能会把关注点放在四个步骤以及每一个步骤涉及的一些基本概念上,例如:筛选获取目的基因的常用方法,鉴定目的基因是否成功导入的方法等等。这当然没错,本题中的(1)(3)考察还是教材上的基础知识。
但从本题的备考来看,这可能不够。教学中还得适当的引入一些近些年的(本题改编自24年的论文)、学生较为陌生的(合成生物学的情境对学生来说是很陌生的)、但又与生活息息相关的基因工程应用(纺织、绿色制造与生活密切相关)。例如,在复习植物基因工程时,可以把2000年左右“华农1号”转基因番木瓜的构建过程作为大情境。复习蛋白质工程时,将CAR-T细胞疗法(改造受体细胞)作为大情境。
最后,也不要总期望课堂或题目中讲过的内容,在高考中就一定能遇到相似的情境。我们要相信,如果我们的教学真的能够使学生的核心素养得到提升,那么即使在高考中遇到完全陌生的情境,学生也依然能够基于自身的素养水平完成作答。
下一期推送会更新对争议较大的19题(植物激素)的文献溯源及点评,敬请期待。
该题改编自2024年发表在Nature Biotechnology的一篇文献:

用工程酪氨酸酶表达的产纤维素细菌培养的自着色纺织品
环境问题正在推动人们对由微生物和真菌来源生产的后石油合成纺织品的兴趣。细菌纤维素(BC)是一种很有前途的可持续皮革替代品,因为它的材料特性,低基础设施需求和可生物降解性。然而,对于像BC这样的替代纺织品来说,要想完全可持续,除了替代生产方法外,还需要开发替代染色纺织品的方法。为了解决这个问题,我们对Komagataeibacter rhaeticus进行基因工程改造,以创造一种能够生长自着色BC的菌株。重组酪氨酸酶在细菌中进行黑色素生物合成,实现了对材料使用坚固的深黑色。黑化BC的生产可以扩大规模,用于原型时装产品的构建,我们通过对纤维素生产细菌基因表达的光遗传学模式,说明了将工程自色素沉积与合成生物学工具相结合的潜力。通过这项研究,我们证明了将基因工程与当前和未来的纺织品生物制造方法相结合,有可能创造出一类新的纺织品。
纺织和皮革工业对环境产生影响,包括农业生产和工业加工产生的温室气体排放、鞣制和染色造成的水污染以及合成纤维脱落造成的微塑料污染。为了减少这个行业的影响,新的可持续生物材料正在商业开发中。这些包括菌丝体和植物纤维皮革替代品。这些努力是将生物生产与工程和化学加工相结合,将这些天然生物材料提炼成可替代纺织品的成功结果。然而,该行业尚未采用这些材料生产生物的基因工程来利用生物系统使用的可持续方法来增强生物材料的物理和美学特性。工程生物材料(ELMs)领域使用合成生物学的工具在DNA水平上对活细胞进行重编程,以构建新的或增强的生物材料,用于特定的应用。细菌纤维素(BC)是一种很有前途的天然生物材料,由革兰氏阴性菌属komagataeibacter细菌最有效地生产。在富含碳的培养基中,这些细菌聚合并分泌线性葡萄糖链。
然后,这些链自组装成紧密相连的纤维素纤维网。这种纤维素网被称为膜,它漂浮在空气-水界面,像生物膜一样包裹和保护生长的细胞。对BC的工业兴趣的关键在于,它可以快速、廉价和可持续地生长——一个BC的膜可以在7-14天内生长,产量高(>10 gl- 1),并且可以从腐烂的果汁、甘油和糖蜜等废原料中生长。此外,BC还具有高抗拉强度、高保水能力和高纯度等先进的材料性能。这些特性引起了人们对BC在高端声学设备、电池隔膜和伤口愈合中使用的兴趣。对于那些寻求可持续纺织品生产方法的设计和时尚人士来说,BC的易于生长也使其成为一种有吸引力的原型生物材料。通过可培养的低风险细菌生产BC也使那些寻求利用合成生物学对其进行基因改造的人能够接触到BC。因此,BC代表了ELM研究的理想“白板”。
BC ELM的研究重点是Komagataeibacter和其他生物(如酿酒酵母)的基因工程,这些生物可以与Komagataeibacter共培养。加入酿酒酵母的使用使得能够感知化学和光刺激并作出反应的膜得以产生。为了方便Komagataeibacter的基因工程,建立了一个模块化的基因克隆工具包,即Komagataeibacter tool kit (KTK),并以Komagataeibacter rhaeticus为载体进行了表征。这个合成生物学工具箱的贡献包括选择模块化的DNA部分,如组成和诱导启动子,载体和荧光标记。
这些部分已被用来制造经过改造的komagataeibacterium,这种细菌可以生产替代聚合物,如几丁质、透明质酸和卷曲纤维。此外,工程的多细胞通信已经在细胞膜上建立起来,通过使用群体感应分子在雷氏克雷伯氏细胞之间进行细胞间的信号传导。然而,尽管取得了这些成就,基因工程尚未用于进一步发展BC作为纺织品和时装的可持续生物材料。
自然界中的生物材料,如头发和皮肤,使用合成细胞产生色素,以低影响的可持续方式在原位为生物材料着色。织物材料的工业着色、纺织染色中使用的平行过程需要化学反应,并且对环境具有高度破坏性。受天然色素生产的启发,我们开始设计一种自着色BC材料,通过基因工程的K. rhaeticus。黑色染料是世界上消耗最多的染料之一,也是最难用可持续染色方法再生的染料之一。我们决定设计黑素的生物合成真黑素,进入雷蒂库斯。真黑素是一种在生物王国中普遍存在的色素,在高温和长时间内保持稳定。至关重要的是,真黑素具有低水溶性,这是许多常见染料(如靛蓝)所共有的特性,有助于颜料的色牢度。此外,真黑素还具有其他一些有趣的特性,如导电性、宽频光和紫外线吸收以及对电离辐射的保护。
我们在这里证明,从红毛蚶中生产的色素纤维素可以大量生产,用于时装产品的原型制作。此外,我们通过生长BC膜中基因表达的光遗传学模式说明了将黑色素生物合成与其他合成生物学工具相结合的潜力。
结果
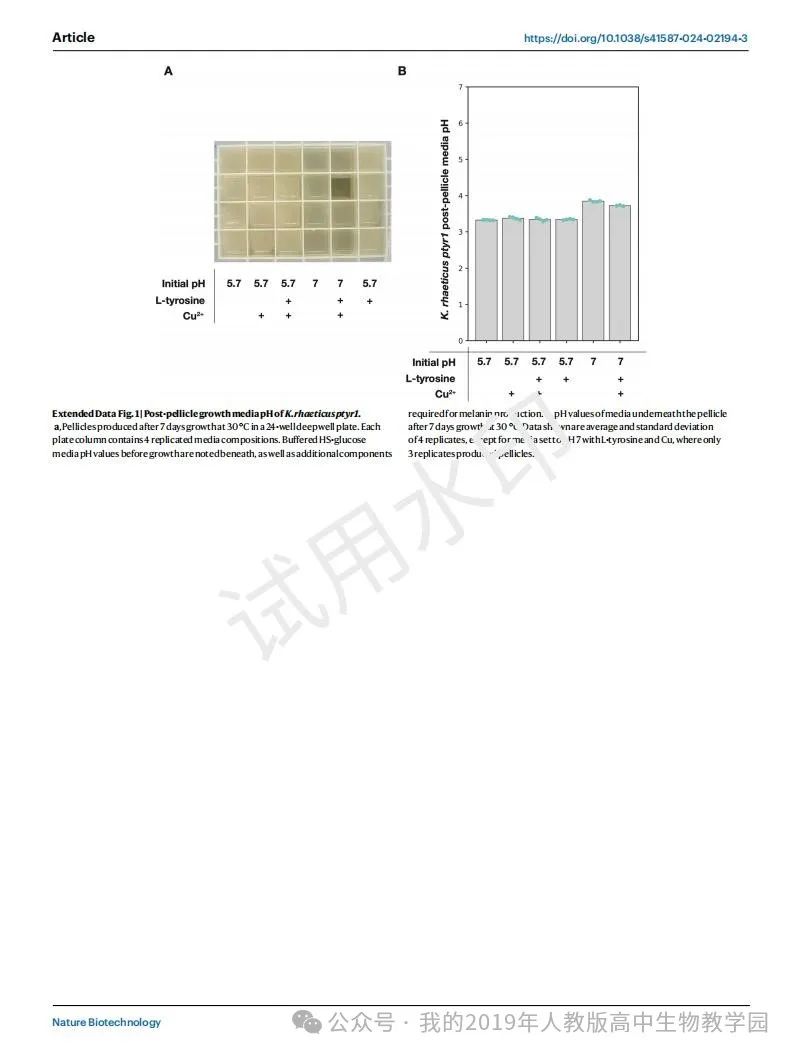
重组生产真黑素已经在大肠杆菌和原生弧菌中得到证实,在生物修复和生物电子学等多种应用中得到应用。细菌生产真黑素只需要一种酶(酪氨酸酶)催化l-酪氨酸氧化为多巴醌,这是真黑素合成的限速步骤。在含氧和温带条件下,多巴醌通过几个步骤自发转化为真黑素(图1a)。已经在重组环境下测试的原核酪氨酸酶有:来自etli根瘤菌的MelA和来自巨芽孢杆菌的Tyr1。我们决定在这项研究中专注于Tyr1,因为它的体积更小,并且在非模式生物中得到了证实。使用我们的KTK系统进行模块化克隆,我们创建了以下两个组成型的rhaeticus Tyr1表达菌株:基于质粒的rhaeticus ptyr1和染色体整合的rhaeticus ctyr1(图1b)。两个菌株在tyr1编码序列周围使用相同的上游和下游DNA部分。tyr1上游的两个构建启动子都是合成组成启动子pJ23104,该启动子在之前的研究中被发现在雷氏K. haeticus26中具有最强的表达强度。
Tyr1的黑色素合成对pH敏感,仅在pH值高于7时才容易发生。这与K的增长相冲突。K.Rhaeticus,作为一种醋酸细菌,在生长过程中通过产生葡萄糖酸和乙酸等有机酸来使培养基酸化。事实上,我们发现在Hestrin-Schramm葡萄糖(hes -glucose)缓冲至pH 5.7并加入生成真黑色素所需的底物和辅助因子(0.5 gl- 1酪氨酸和10 μM Cu2SO4)的培养基中生长的rhaeticus ptyr1膜在生长过程中没有色素沉着(扩展数据图1a)。我们测量了生长培养基在生产膜后的酸化情况,结果表明,即使初始培养基的pH被缓冲到pH 7,培养基的pH也已降至pH 4以下(扩展数据图1b)。这些结果表明,我们需要将膜生长与真黑色素的产生分离开来。
因此,我们决定采用两步法从红僵菌中产生黑色素。第一步是在正常生长条件下培养表达tyr1的rhaeticus,第二步是去除废培养基并用含有合成黑色素所需试剂的缓冲溶液代替(图1c)。对于我们称为黑色素发育缓冲液的缓冲溶液,我们选择使用PBS,缓冲至pH 7.4,含有0.5 g l- 1酪氨酸和10 μM Cu2SO4。然后,我们在两个表达tyr1的菌株上测试了这种方法。为了量化真黑素的产生,我们检测了在摇动条件下生长的雷氏克雷伯细胞,培养基中添加了纤维素酶,以防止膜的形成。我们测量了在OD405下黑色素发育缓冲液中12小时的真黑色素生成(图1d)。在发育缓冲液中,两种表达tyr1的雷蒂库菌都能产生真黑素。综合酪氨酸酶菌株rhaeticus ctyr1的每个初始细胞黑色素生成率(0.48±0.03 OD405/OD600/h)高于基于质粒的rhaeticus ptyr1(0.35±0.02 OD405/OD600/h);图1 e)。我们还使用相同的实验方法来分析改变黑色素发育缓冲液的pH、盐浓度、氧化态(II)金属离子和铜(II)浓度对黑色素生成的影响(扩展数据图2a-h)。有趣的是,我们发现碱性缓冲条件(>pH 8)导致真黑素的生产比中性条件更快。然而,碱性缓冲条件下的黑色素生成速度也迅速减慢,导致黑色素积累总体低于pH 8。这一结果与之前对Tyr1的体外研究相矛盾,该研究认为Tyr1的最佳pH值为7。然而,同样的研究也表明,当pH值高于7.5时,左旋多巴可以自发地转化为多巴胺。这里看到的体内结果可能反映了细胞内积累的l-DOPA的自发转化,这可能表明,尽管缺乏可见的黑色素生成,但在rhaeticus生长阶段,Tyr1具有一定水平的单酚酶活性(l-酪氨酸到l-DOPA)。
然后,我们观察了雷氏克雷伯氏细胞在暴露于黑色素发育缓冲液后是如何变化的。通过光镜观察,我们发现暴露于发育缓冲液中的rhaeticus ptyr1和cytyr1细胞明显变暗,表明真黑色素的产生可能发生在细胞内(图1f)。我们还对rhaeticus ptyr1和Tyr1培养物的上清液进行了黑色素生成实验,发现有和没有l-酪氨酸的上清样品在黑色素积累率上没有显著差异,这表明细胞外Tyr1在摇晃培养物中的存在是最小的(扩展数据图3)。考虑到Tyr1蛋白不含分泌或易位标签,这是意料之中的。然而,由于真黑素的产生需要细胞浸泡在中性pH缓冲液中,这也表明在生长过程中,雷氏克雷伯氏菌的细胞质pH也可能变为酸性。事实上,其他的醋酸细菌也表现出对酸性内部环境的适应性。

图1:酪氨酸酶表达产生真黑素。
a, l-酪氨酸生成真黑素的化学途径。第一步包括l-酪氨酸羟基化成l-多巴,这是由酪氨酸酶催化的,这里作为一种单酚单加氧酶。这一步之后是左旋多巴催化多巴醌,这是由Tyr1的二酚酶活性催化。该途径的其余步骤在氧气存在的情况下自发发生,导致真黑素的产生。
b,以下两种rhaeticus酪氨酸酶表达菌株的遗传结构图:rhaeticus ptyr1和rhaeticus ctyr1。这两种结构使用相同的组成启动子(pJ23104)、RBS (B0034)和终止子(L3S1P00)。rhaeticus ptyr1使用具有pBBR1复制起源的质粒和氯霉素抗性盒。
c,在摇晃条件下生长的雷蒂库菌生产真黑素的两步法。菌株在hs -葡萄糖培养基中培养,用PBS洗涤并重悬以去除废培养基,然后与黑色素发育缓冲液混合。
d,检测tyr1产生菌株的真黑色素产量。每个初始细胞的真黑色素产量是通过测量12小时内的OD405,除以每口井在时间点0的初始OD600来确定的。点表示三个生物重复的平均值。误差条是三个生物复制的标准差。
e,测量开始后50 ~ 170 min,测量每个初始细胞的真黑色素积累梯度,确定每个初始细胞的初始反应速率。条形图显示了每个菌株三次生物重复的平均值,而误差条显示了黑素发育前(mel−)和黑素发育后(mel+)的sd - f光学显微镜图像。单个单元格的放大示例显示为青色轮廓。所示的图像是为每种菌株和治疗所拍摄的四张随机图像的代表。

图2 |利用表达tyr1的rhaeticus产生黑化BC。
a、黑化BC的生产过程包括两个步骤。表达酪氨酸酶的雷氏菌在静态条件下生长以产生膜。一旦生长,将其收集并放置在显影缓冲液中,在30°C至50°C之间搅拌孵育,直到材料达到所需的阴影。
b,图像显示了在30°C下放置在发育缓冲液中的K. rhaeticus ptyr1和K. rhaeticus ctyr1膜在24小时内真黑色素积累的时间变化。
c、薄膜生产可以在标准化的容器中进行,以生产出BC片,从中裁剪出图案片并进行组装。
d,在300 × 200 mm的容器中生长,经过真黑色素发育步骤的A . rhaeticus tyr1被膜。
e,一个最终的钱包原型,由两张压干的黑化BC纸切割和组装而成。
f、薄膜生产也可以发生在异形容器中,生产BC预成型为最终图案片的2D图案。g,在真黑色素发育后,在一个形状容器中生长的金貂鱼ptyr1膜。在膜的周长内看到的金属钉固定着编织的线网,在生长过程中融入到膜中。
h,一种最终的鞋面原型,由黑化形状的BC片和集成的纱线制成,该纱线缠绕在脚形的鞋楦上,并放置在鞋底上。
在证实了表达tyr1的雷氏克雷伯氏细胞能产生真黑素之后,我们下一步想要证明真黑素的产生能有效地使BC色素化。为了做到这一点,我们将同样的两步过程应用于已生长膜的雷蒂库菌ptyr1和ctyr1静态培养物(图2a)。在30°C的发育缓冲液中振荡孵育24小时后,膜的外观从淡黄色变为棕黑色,表明BC的真黑色素色素沉着(图2b)。在对红毛菌ptyr1和ctyr1随时间的视觉变暗进行量化后,在测试条件下,我们发现它们在19小时后达到峰值可见黑暗(扩展数据图4)。此外,通过降低发育缓冲液中的l-酪氨酸浓度,我们可以减缓黑色素产生的速度(扩展数据图5a),使我们能够改变BC膜的着色方式,从而产生一系列棕色色调的物质(扩展数据图5b)。从水合黑色素膜的薄截面切片中,我们观察到黑色素色素沉着在膜的顶部最深(扩展数据图5c)。我们还发现,在培养基和黑色素发育缓冲液中加入0.5 g l-酪氨酸会导致最暗的膜,可能是因为这允许细胞在生长过程中积累l-酪氨酸水平和真黑色素前体。在实验室环境之外使用黑化BC的关键是,这种色素可以通过灭菌而持续存在。我们发现高压蒸汽和乙醇灭菌都能很好地保存色素沉着(扩展数据图5d)。正如预期的那样,通过氧化化合物(如次氯酸钠漂白剂)进行杀菌,导致黑色素色素沉着迅速消失。最后,我们测试了黑化BC对水渍的色牢度——这是一种常见的皮革污渍测试。我们发现黑化BC的颜色非常稳定,没有因水斑而可见的变色(扩展数据图6)。
由于从简单的静态生长培养中产生的材料产量很高,细菌的微生物生产非常适合扩大规模,从而能够以最小的基础设施投资实现构建实际产品所需的细菌数量。这使得BC对工业和家庭工业规模的生产商都很有吸引力,尤其是作为服装和配饰中皮革的素食替代品。考虑到这一点,我们想要证明,我们可以扩大表达tyr1的雷提克雷伯菌的生长,以产生功能上有用的色素BC。为此,我们考虑了BC生产的两种方法。在我们的第一种方法中,我们试图生产标准化的黑化BC片,从中可以切割和组装非织造纺织品图案(即模板)(图2c)。为了帮助BC大规模生长,我们改用含有椰子水、1%乙醇和1%醋酸的生长培养基。该培养基在工业环境中用于培养雷氏克雷伯氏菌并使BC产量最大化48。我们在300 × 200 mm的培养皿中培养雷氏克雷氏菌(K. rhaeticus tyr1),生长10天后,我们收集其膜,并让其产生真黑色素,直到其呈现深黑色(图2d)。黑化的BC片然后用高压灭菌器灭菌,压平并干燥。BC薄片在整个过程中保持其颜色(补充视频1)。然后从这些黑色素薄片中裁剪出一个钱包图案,并用线将图案片缝在一起,制成一个功能正常的黑色素BC钱包(图2e)。
在我们的第二种方法中,我们利用了膜生长遵循空气-水界面的方式,并以与培养容器相同的形状生长(图2f)。利用雷氏克雷伯氏菌ptyr1,我们在一个定制的培养容器中培养了一个膜,形状像鞋面图案。这个培养容器中有一个织布机状的装置,上面有一个由Lyocell (TENCEL)线串成的网络,位于空气-水界面,使这些线能够融入正在生长的海蝇ptyr1细胞膜中。生长14天后,将终膜和装置从培养基中取出,置于发育缓冲液中。在30℃下轻摇48小时后,膜呈现深黑色(图2g)。然后用乙醇浴对薄膜进行灭菌,并将其浸泡在5%甘油溶液中,然后从仪器中取出,包裹在环氧树脂鞋楦(即脚形模具)周围并使其干燥(图2h)。
黑化BC,以及标准的非转基因BC材料,仍然需要额外的加工和添加剂,以满足当前替代皮革材料性能的期望。尽管如此,我们相信鞋面和钱包都表明,我们的工程菌株可以生长和自我色素,其规模足以生产出原型时装作品,这可以展示黑色素化BC的可能形式和美学。这些作品也展示了科学家和设计师在追求创造新的elm时合作的积极成果。作为新型生物材料纺织品的使用者,设计师在展示和宣传新材料的特性方面发挥着关键作用,并且可以就任何限制以及如何改进材料向科学家提供建设性的反馈,特别是为了使其寿命结束时变得更加可持续。
由K.rhaeticus ptyr1产生的黑化纤维素酶样本被积极用作42个月的样品,并在整个过程中保持其色素沉着(图3a),表明颜色随时间的推移具有弹性。除了颜色,我们很想知道真黑素的产生是如何影响BC的其他物质特性的。为了研究这一点,我们首先使用扫描电子显微镜(SEM)检查真黑素是否改变了BC表面。
我们比较了黑化和未黑化的雷蒂库菌ptyr1膜的上下表面以及截面(图3b)。扫描电镜图像显示,黑化和非黑化膜之间的结构差异很小。顶部表面图像上的不均匀表面形貌归因于剩余的嵌入细胞,而底部表面的形貌则更光滑。此外,黑化和未黑化的ptyr1膜的横截面显示BC纳米纤维网络的孔隙度差异很小。为了进一步研究黑化纤维素的表面材料特性,我们使用静态无根滴法进行润湿性测试(图3c)。利用从黑僵菌ptyr1中生长的膜,我们观察到黑化后的膜具有更高的表面润湿性,平均接触角为28°,而未黑化的膜为47°。
BC对行业最具吸引力的特点之一是其高抗拉强度;因此,了解黑色素是干扰还是增强BC纳米纤维网络的强度是很重要的。我们使用黑化和未黑化的薄膜进行拉伸测试。为了保持一致性,我们准备了一组配对的BC样品,将每个生长的膜分成两半,并在每个膜的一半中产生真黑色素。两半都被热压,以巩固BC纳米纤维网络成干燥的BC片(图3d)。未黑化和黑化膜的平均抗拉强度分别为91 MPa和105 MPa(图3e)。对于杨氏模量,未黑化和黑化的膜分别为13.7 GPa和13.9 GPa(图3f)。对于断裂应变,未黑化和黑化样品的值分别为1.02%和0.98%(图3g)。所有测试样品的材料性能均在BC抗拉强度70-300 MPa和杨氏模量5-17 GPa的预期范围内。配对t检验表明,黑化和未黑化膜的干燥BC在拉伸材料性能上没有显著的统计学差异。
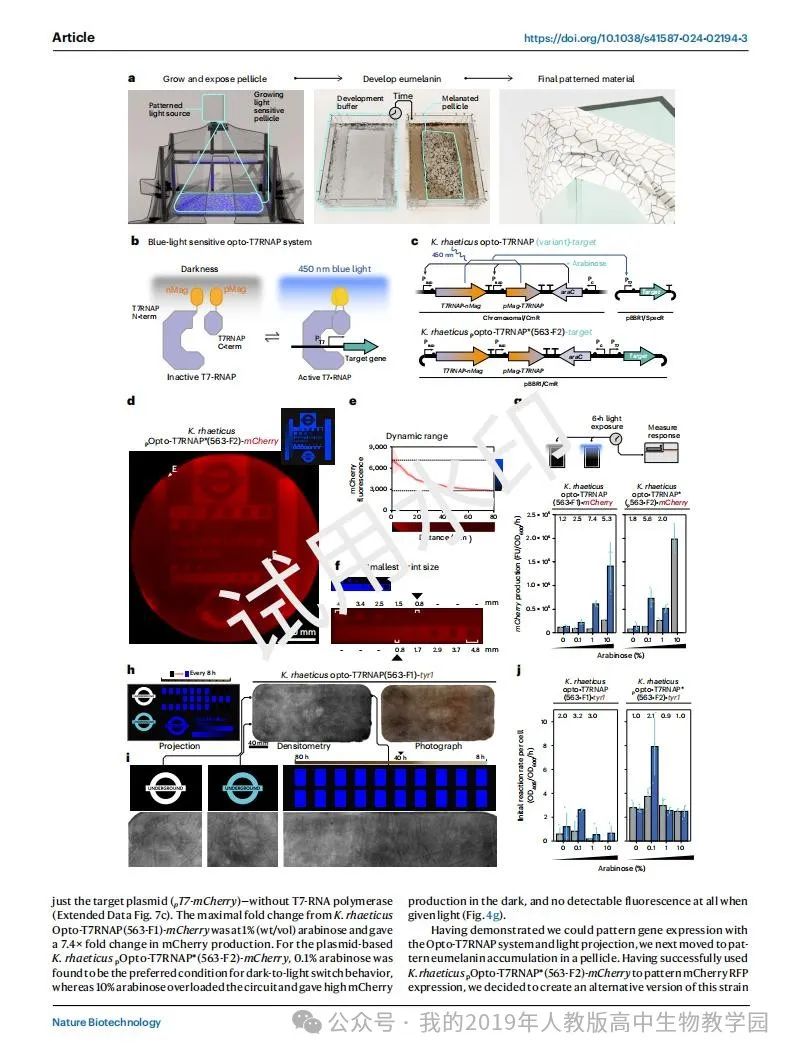
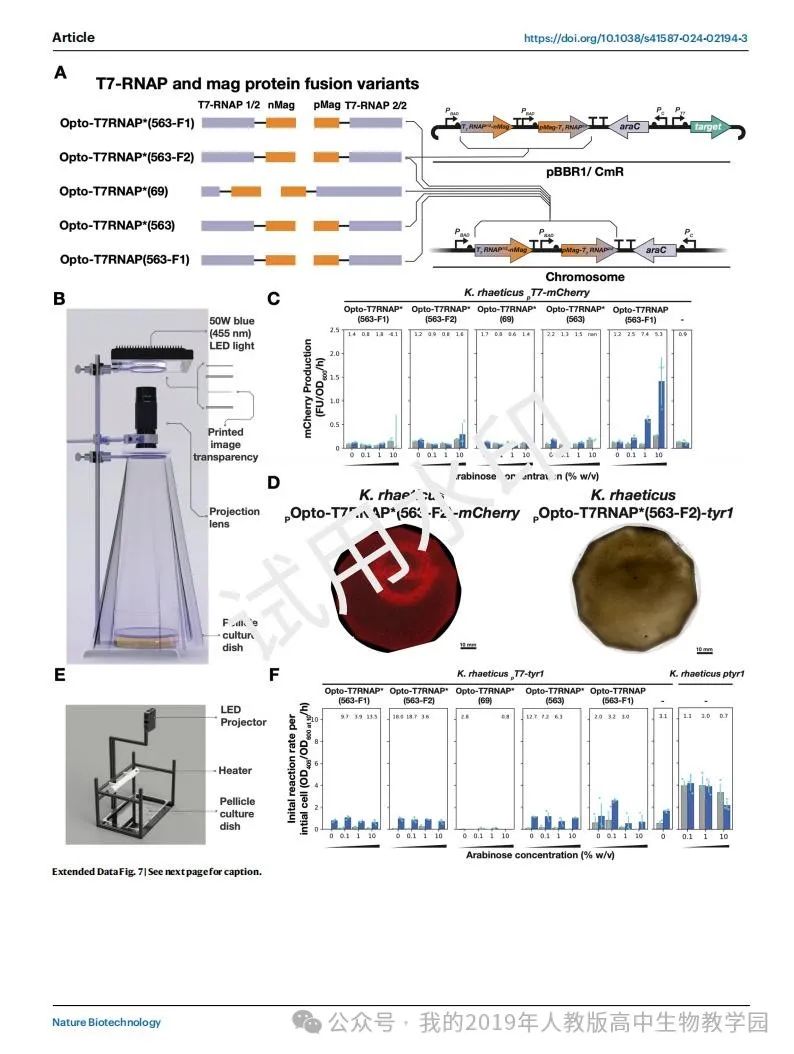
除了上色,纺织品加工还包括给纺织品打图案。为了展示基因工程的自着色BC无纺布的能力,我们开始建立基因表达的空间控制。我们确定最灵活和精确的基因表达模式的方法是通过工程光遗传学,并提出了用光制造模式BC的程序(图4a)。为了设计一种光敏的雷提库菌菌株,我们选择了采用最初由Baumschlager等人设计用于大肠杆菌的蓝光敏感T7-RNA聚合酶(Opto-T7RNAP)系统(图4b)。我们推测,Opto-T7RNAP系统将是最简单的光遗传系统之一,可在非模式生物中实现,如金鼠50。这一观点是基于T7-RNA聚合酶转录的正交性,缺乏膜结合的光敏成分,以及使用普遍存在的黄素腺苷二核苷酸(FAD)作为发色团,从而消除了对异源发色团生物合成基因的需要。
为了实现Opto-T7RNAP系统,我们测试了两种必要DNA部分的排列。我们选择参考文献49中提到的光照和暗态之间fold变化最大的变体(Opto-T7RNAP*(563-F2))作为基于质粒的变体,将光敏T7RNAP基因和光调节靶基因置于同一质粒上(扩展数据图7a)。为了测试Baumschlager等人49生产的Opto-T7RNAP系统的变异,我们将光敏T7RNAP变异基因整合到雷舌鼠染色体中。这些K。然后用编码靶基因的单独质粒转化rhaeticus T7RNAP变异菌株(图4c)。在这两种安排中,两个Opto-T7RNAP光敏基因都受到PBAD启动子的调控,PBAD启动子先前已被证明在2% (wt/vol)浓度的阿拉伯糖25诱导下在雷氏乳杆菌中起作用。在染色体和质粒排列中,阿拉伯糖调节基因araC位于整合的两个Opto-T7RNAP光敏基因的下游。
然后,我们测试了是否可以使用Opto-T7RNAP系统对雷蒂库菌的基因表达进行模式化。为了简化这一过程,我们首先将目标基因作为产生红色荧光蛋白(RFP) mCherry的荧光报告基因。我们构建了一个投影装置,可以在培养液生长时将图像投影到培养液表面(扩展数据图7b)。我们在该装置的培养容器中接种rhaeticus pOpto-T7RNAP*(563-F2)-mCherry,一旦形成薄的膜,我们将这个新生的膜暴露在投影图像中72小时。收集到的细胞膜显示mCherry的成功表达模式(图4d)。在最不暴露部分和最暴露部分的膜膜荧光变化为2.54倍(图4e)。值得注意的是,我们还发现我们可以视觉模式的最小区域为0.8 mm2,这给了我们一个方法分辨率的工作估计(图4f)。

图3 |黑化BC的性质。
a,黑化的(mel+)和未黑化的(mel−)样本,来自相同的原始金貂鱼ptyr1细胞膜。这些样品被晒干后用作展示品。
b, mel+和mel - rhaeticus ptyr1 BC的SEM。顶面和底面分别属于面向空气和面向介质的膜面。所示的图像是每种情况下至少五个图像的代表。
c、用无柄滴法测定了雷蝇ptyr1 mel−(米色)和mel+(黑色)膜上的接触角。非配对t检验结果的值P < 0.005,误差条表示8梅尔-和9梅尔+跌落测量的标准差。
mel+和mel -膜的代表性水滴形状如图所示。
d,对黑化和未黑化的BC片进行了对比拉伸试验,使用的BC片是由相同的雷氏库菌ptyr1膜的一半制备的。给出了mel -和mel+的BC断裂的代表性图像,以及mel+和mel -三个生物重复的技术重复的应力-应变曲线。
e - g, mel+和mel -膜的抗拉强度(e),杨氏模量(f)和断裂应变(g) - mel+和mel - BC之间的配对t检验的P值分别为0.17,0.92和0.85。误差条显示来自三个生物复制的sd,每个生物复制是三个或更多技术复制的平均值。
然后,我们开始了解光传感器表达在红齿K. rhaeticus的最佳水平,以最大限度地提高光和暗状态之间的动态范围。我们通过测量暴露于一系列阿拉伯糖浓度的雷氏克雷伯氏菌Opto-T7RANP变异在微滴板中平行生长时的蓝光响应来研究这一点。在这些实验中,我们测试了最初由Baumschlager等人创建的Opto-T7RANP系统的五种变体,这些变体被放置在雷氏克雷伯氏菌的染色体上(扩展数据图7c)。在评估蓝光照射6小时后的mCherry表达反应后,我们发现只有一个整合变体(k.r eticus Opto-T7RNAP(563-F1)-mCherry)的mCherry表达水平高于基线(k.r eticus pT7-mCherry),阴性对照菌株只含有目标质粒(pT7-mCherry),没有T7-RNA聚合酶(扩展数据图7c)。K. rhaeticus Opto-T7RNAP(563-F1)-mCherry在1% (wt/vol)阿拉伯糖浓度时最大的折叠变化,使mCherry产量变化7.4倍。对于基于质粒的rhaeticus pOpto-T7RNAP*(563-F2)-mCherry, 0.1%阿拉伯糖被发现是暗到光切换行为的首选条件,而10%阿拉伯糖使电路过载,在黑暗中产生高mCherry,在光照下根本没有可检测到的荧光(图4)。

图4 |K.rhaeticus的功能光遗传学。
a,提出了通过光遗传学制造图案化BC的程序。
b, Opto-T7RNAP系统使用分裂的T7-RNA聚合酶,通过与光敏磁铁蛋白融合使蓝光活化。
c,金貂鱼光遗传菌株的遗传排列。用阿拉伯糖诱导分裂的T7RNAP基因表达。
d,蓝色光暴露湿的rhaeticus pOpto-T7RNAP*(563-F2)-mCherry膜(直径= 150 mm)顶部表面的红色荧光扫描。右上方的图像显示了生长过程中投影到细胞膜上的图像。所示的膜是两个有图案的膜重复的代表。
e、投影图像的右侧包含一个渐变带,从最小到最大的光线通过。数据显示红色荧光的强度在膜相对于这个渐变条。每个水平切片的像素强度sd用粉红色表示。
黑色虚线表示未暴露的膜区强度。
f,暴露在外的膜上最小的投影标记。
g,不同阿拉伯糖百分比(wt/vol)浓度下mCherry靶基因光遗传学构建体的表征。条形图(蓝色,曝光,灰色,未曝光)显示经OD600归一化6小时后红色荧光的平均增加。误差条显示s.d。在三个生物复制中。除了生长不良的情况外,暴露和未暴露的细胞之间的倍数差异如上所示。
h,投影视频与真黑素发育后得到的湿K. rhaeticus opt - t7rnap (563-F1)-tyr1膜的比较(尺寸= 300 × 170 mm)。投影视频顶部的矩形(黑色和蓝色)被定时显示,以帮助计算最小光照时间。给出了膜顶表面的密度扫描图和照片。
i, K的密度扫描的放大切片。Opto-T7RNAP(563-F1)-tyr1膜。黑色三角形指向第六个矩形,表示需要曝光40小时。
j,不同阿拉伯糖诱导下tyr1的光遗传学结构表征。柱状图(蓝色,暴露,灰色,未暴露)显示在OD405测量的真黑素产生的初始(0 - 100分钟)反应速率的三个生物重复的平均值和标准差,归一化到OD600在时间点0的初始细胞数量。
在证明了我们可以用Opto-T7RNAP系统和光投射来模拟基因表达之后,我们下一步要模拟真黑色素在细胞膜中的积累。在成功使用K. rhaeticus pOpto-T7RNAP*(563-F2)-mCherry对mCherry RFP表达进行模式分析后,我们决定创建该菌株的替代版本,将mCherry编码序列直接替换为tyr1编码序列,从而创建K. rhaeticus pOpto-T7RNAP*(563-F2)-tyr1。然而,当与mCherry版本并行测试时,我们发现tyr1版本具有如此高水平的背景真黑素产生,以至于它掩盖了可见的真黑素积累模式(扩展数据图7d)。因此,我们改用另一种Opto-T7RNAP变体,该变体在使用mCherry靶基因- k的摇晃条件实验中也表现出明显的蓝光反应。rhaeticus Opto-T7RNAP(563-F1)进一步测试我们是否可以模拟真黑色素的积累。
在测试之前,我们设计了一个要投影的动态图像和一个新的商业投影仪投影设置,允许我们在曝光期间改变图像,从而测量所需的曝光时间,以产生可观的真黑色素响应(扩展数据图7e)。我们使用K. rhaeticus Opto-T7RNAP(563-F1)-tyr1,在该装置中培养了一个BC膜,一旦生长,我们将膜暴露在80小时的投影中(图4小时)。然后,我们将暴露的细胞膜放入发育缓冲液中,在30°C下孵育48小时,此时我们可以观察到真黑色素积累的大致模式(图4i)。不幸的是,尽管在某些地方显示出一些图案的证据,但高度的背景真黑素积累也使这种图案的尝试难以破译。然而,我们可以从动态模式中确定,当K. rhaeticus Opto-T7RNAP(563-F1)-tyr1产生真黑色素时,至少需要40小时才能观察到可见的真黑色素积累。
最后,我们评估了其他Opto-T7RNAP变体以tyr1作为其靶基因时的表现。我们使用了与之前类似的平行微滴板方法,但现在在OD405测量每个初始细胞随时间的真黑素积累(图4j)。结果表明,rhaeticus pOpto-T7RNAP*(563-F2)-tyr1需要0.1%的阿拉伯糖才能正常工作,就像mCherry作为靶标时一样(图4)。然而,现在在这种条件下和所有其他测试中,即使在没有光诱导的情况下,也观察到高真黑素的产生率。这一发现与在试验膜中看到的高背景色素沉着一致。在染色体整合DNA的菌株中,K. rhaeticus Opto-T7RNAP(563-F1)-tyr1对蓝光的响应显示出最高的真黑素产量,同样需要0.1%的阿拉伯糖来调节。该菌株的明暗状态之间的折叠变化低于测试的其他两种变体,这两种变体的折叠变化都大于10倍(扩展数据图7F)。然而,这些菌株不适合色素沉着,因为它们的真黑色素产量要低得多。总的来说,虽然我们可以使用Opto-T7RNAP系统显示蓝光可以调节真黑素的产生,但真黑素在细胞膜中积累的准确模式仍有待优化。
我们使用Opto-T7RNAP对真黑色素积累进行图图化的主要限制是高水平的背景色素沉着和对蓝光响应的受限折叠变化。这两个因素严重降低了系统的动态范围。其他可能也降低了膜上模式定义的因素包括Tyr1酶或细胞中l-DOPA的泄漏(例如通过细胞裂解),或者Tyr1的表达导致蓝光暴露区域细胞生长和密度降低。大多数这些因素可以通过降低目标质粒的背景tyr1表达来解决,但这种背景表达的确切来源目前尚不清楚。优化雷蒂库菌中Opto-T7RNAP基因的排列,有望使我们接近大肠杆菌中该系统的动态范围。我们预计,在不久的将来,提高Opto-T7RNAP系统在雷蒂库菌中的性能,并开发替代靶基因,将产生更先进的BC生物材料。
讨论
在这里,我们将基因工程应用于一种生产材料的细菌,目的是制造色素产品。我们已经证明,酪氨酸酶表达的克雷伯氏菌是熟练产生高度着色的BC;工程红僵菌的生长可以按比例产生有用数量的色素BC,并且这种色素沉着是稳定的。这项研究证明了利用基因工程设计和构建菌株的价值,这些菌株旨在培养具有所需性能的材料;在这种情况下,用一种选定的颜色生长在材料中,而不是稍后通过工业化学染色过程添加到材料中。
我们的研究只是黑化BC发展的第一步。采用两步法生产真黑素增加了黑化BC所需的水量,这可能限制其可持续性。潜在地,这种额外的水的使用可以通过开发缓冲液的再利用和再循环,或在膜生产后将缓冲成分直接添加到生长介质中来减少。然而,最有效的方法可能是调整真黑素的生产,以发生在由红僵菌产生的酸性条件下。这种方法需要对真黑素化学途径进行评估,以确定对低pH值敏感的来源,并对天然黑色素生物合成机制进行调查,以确定耐酸途径。耐酸酪氨酸酶的鉴定或定向进化是一个特别有前途的途径,可能使未来的一步过程成为可能。
我们相信,黑化BC的生产可以在工业环境中进一步扩大规模。虽然我们已经证明了Tyr1在雷氏克雷伯菌中起作用,但许多其他komagataebacter菌株被用于工业BC生产,并且每种菌株的培养条件各不相同,以最大化产量。因此,为了增加我们研究的灵活性,下一步应该在其他Komagataeibacter物种中进行tyr1表达的基因工程研究。在工业层面,重要的是,黑化BC要接受更严格的色牢度行业标准测试,比如在紫外线和可见光下耐摩擦和褪色。
此外,黑化BC的可持续性证书也应通过全生命周期分析进行评估。黑化BC还将面临已经限制BC作为非织造纺织品使用的挑战。主要是,BC的高亲水性要求在纺织加工阶段具有显著的防水性能。各种疏水涂层已被用于用合成或天然化合物对纤维素基材料进行防水,这种技术已被证明通过用植物来源的蜡和油涂层来降低BC的亲水性。然而,这一过程也可能通过Komagataeibacter的基因工程重新考虑,要么改变生长的BC结构,要么通过生物合成一层疏水化合物。此外,BC材料的灭菌,特别是在高压蒸汽灭菌的情况下,需要考虑大量的能量需求。
最后,还有其他途径来探索基因工程的方法,在这里演示,以产生其他色素分子从雷蒂库。酪氨酸酶产生的左旋多巴在有半胱氨酸存在的情况下,可转化为红色素、褐黑素的形成,进一步的研究将扩大黑化BC物质可能的色度范围[52]。其他不溶性色素分子的生物合成也可以进行。最明显的是靛蓝染料的生产,如靛蓝和提尔紫,其生物合成已经在大肠杆菌中得到证实。事实上,由于在遗传水平上混合色素生产的潜力以及通过不同色素生产菌株的共培养,仅一小部分色素的工程生物合成可以提供额外的BC色调范围。






















论文下载链接:https://pan.baidu.com/s/1tWqKSiUViHvYct2F4FXwVQ?pwd=xi3k 提取码:xi3k
原题呈现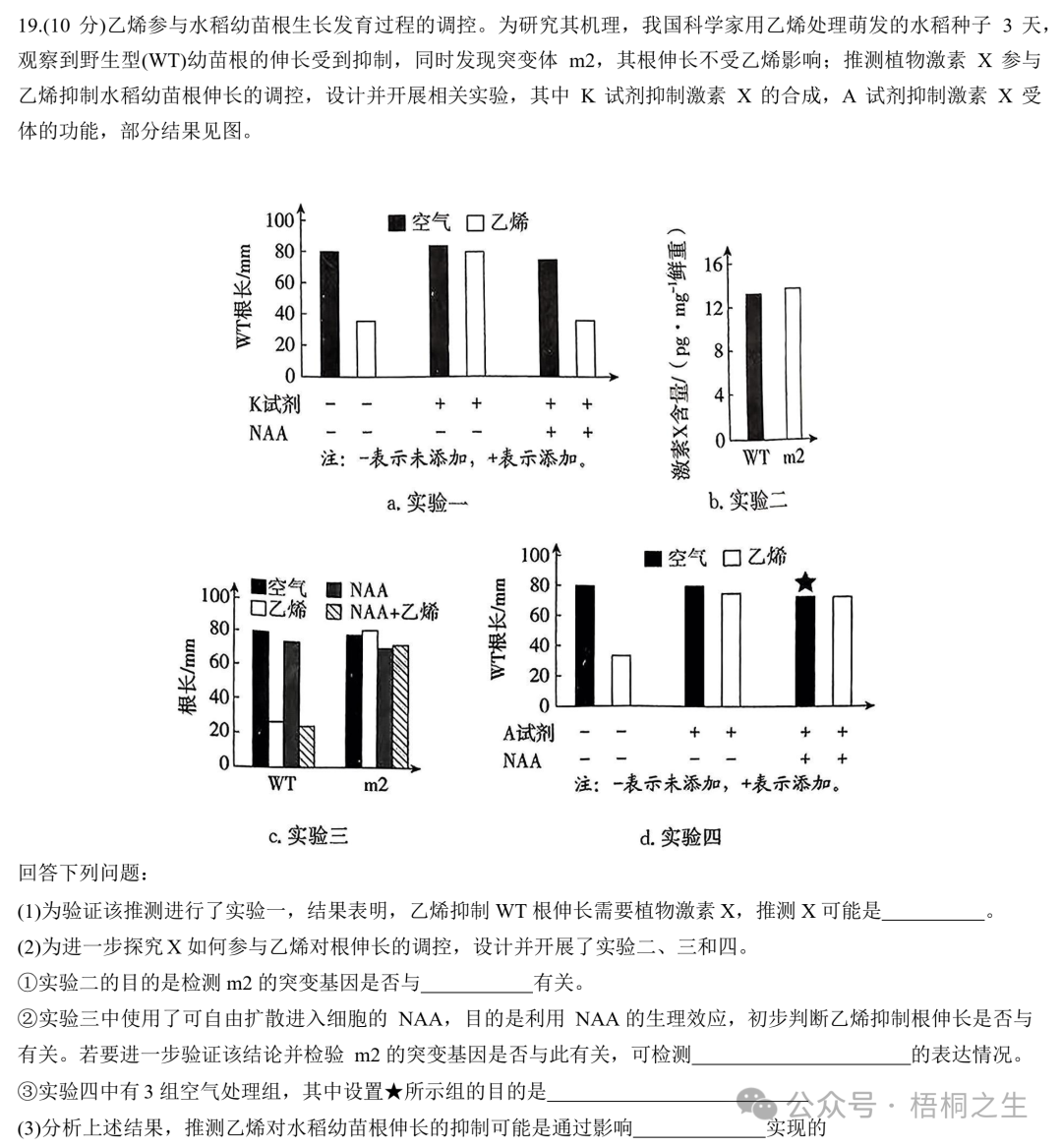
网传版本的答案:
(1)生长素(1分)
(2)①生长素的合成(2分)
②生长素的运输(2分) 载体蛋白的基因(1分) 作为实验组,③检测A试剂和NAA对根生长的影响;与乙烯组作对照(共2分)
(3)生长素(或植物激素X或NAA)的信号(或信号转导)(2分)
根据19题提供的四个实验来看,这节选自近年发表在PNAS的一篇文献,网传来自Plant Cell有误。由于原题干删去了一些关键信息,我不想根据有限的信息硬生生搬出一套牵强的解释。所以,对19题的分析完全参照原文献的写作顺序,重点分析①科学家要解决的科学问题及②实验1-4之间的逻辑关系。
实验1原文献中首先提到,在拟南芥中发现生长素功能依赖的蛋白质突变后,会导致无法观测到乙烯反应,这说明乙烯反应与生长素有关。于是,作者提出了第一个问题:拟南芥中如此,那么水稻根系的乙烯反应(乙烯反应这个说法遵照原文,指乙烯可诱导水稻幼苗根伸长受到抑制)是否也需要生长素参与?由此展开了实验1。
4个实验中实验1相对好理解,通过运用减法和加法原理来探究生长素在乙烯反应中的作用。减法原理对应施用生长素合成抑制剂,结果观察到乙烯反应的消失。加法原理则对应在施用生长素合成抑制剂的基础上,继续补充NAA,重新观测到乙烯反应的出现。一减一加证明了水稻根系的乙烯反应确实需要生长素参与,实验1对应原题第(1)问。
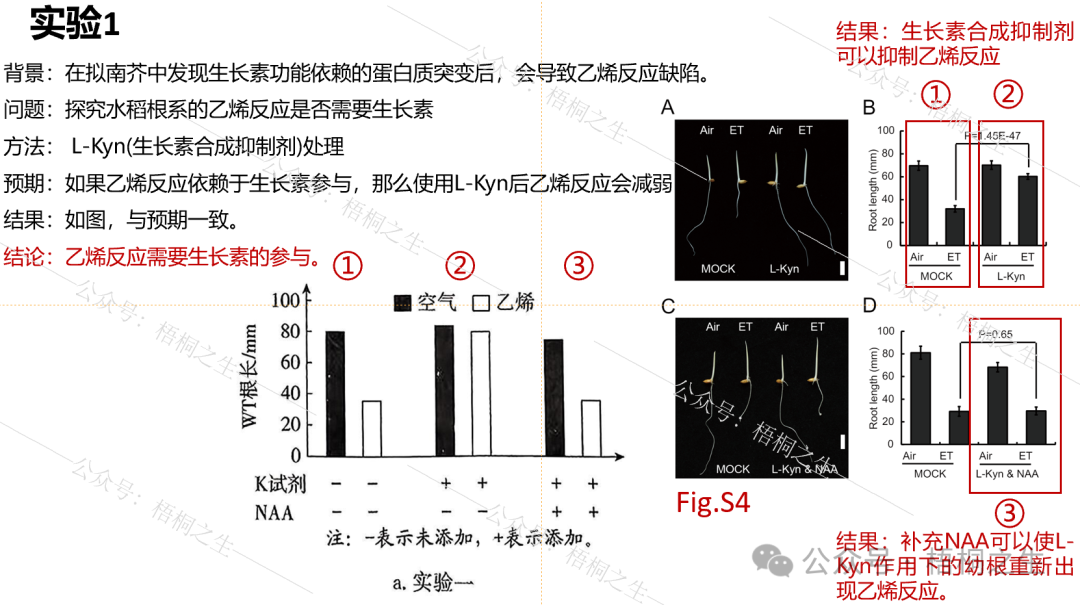
MOCK表示空白对照,L-Kyn即K试剂。这里用①②③把题干中的分组与文献中的分组对应起来。
这里忍不住吐槽一下(1)的设问:
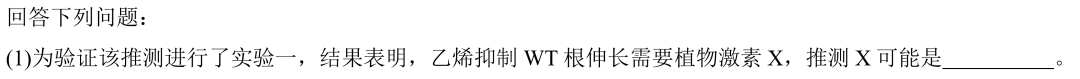
如果命题人本意考察一点科学思维+基础知识,希望考生通过分析实验1的柱状图,发现使用NAA后可重新出现乙烯反应,而NAA又作为生长素类似物,由此推测激素X是生长素。那么为什么要把实验1的结果直接在题干中呈现出来,浪费考生时间去分析实验1?
如果命题人是希望考生看到NAA三个字母就想起X是生长素,那这就纯属记忆层次的考察。既然考察的是死记,那又何必展示实验1,甚至在题干中把实验1的结果再复述一遍,直接删去实验1,然后把第(1)问换成"实验中所用的NAA是的类似物"不是更简单么?优秀的学生在解题时内心一定是崩溃的,NAA不就是生长素类似物么,太简单了!不对,好像没有用上实验1柱状图的信息啊,容我再思考思考!咦…我分析出来了,但是实验1的结果怎么在题干中直接告诉我了??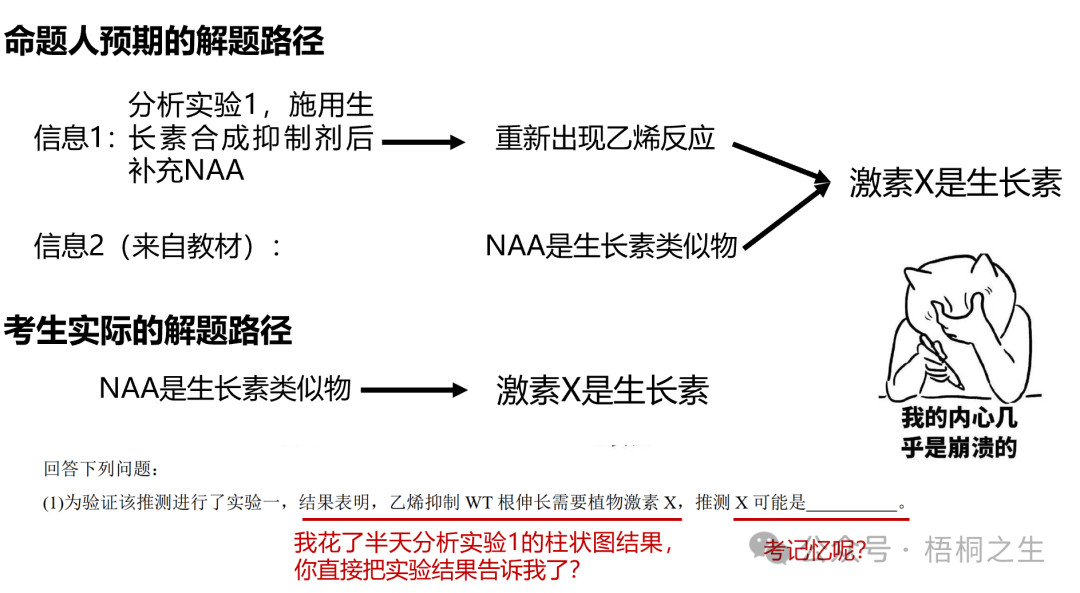
在实验1基础上,作者进一步好奇:既然生长素参与乙烯反应,那么生长素信号通路对于触发乙烯反应来说是必要的吗?或者换句话说,水稻根系触发乙烯反应是否一定要经过生长素信号通路来实现?为了完成这一必要性论证,可以通过减法原则来实现(使用受体抑制剂)。
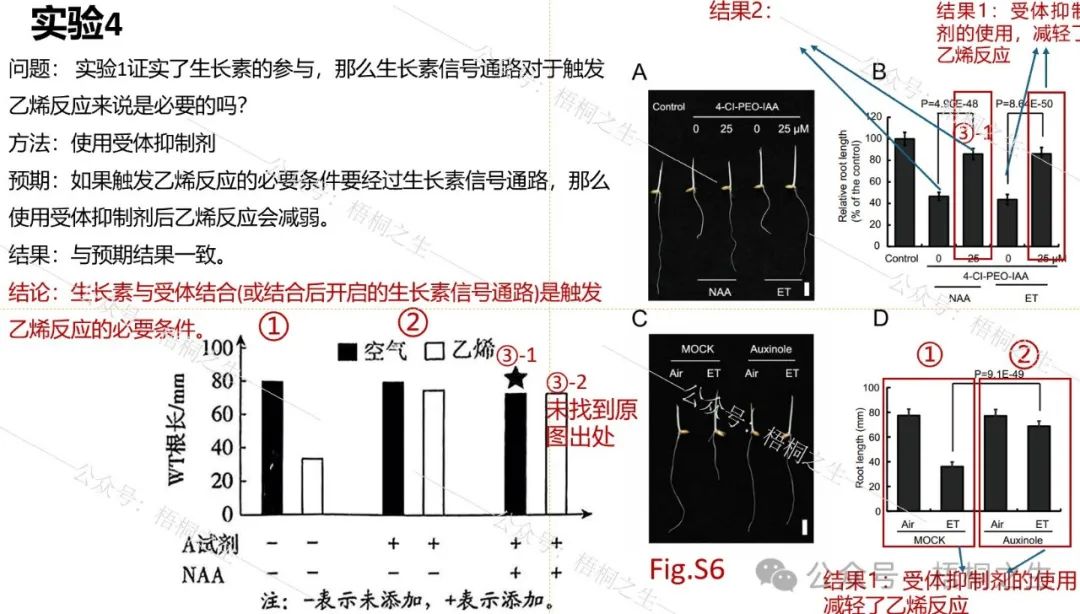
Auxinole和4-CI-PEO-IAA为生长素受体抑制剂,即题中的A试剂。这里用①②③把题干中的分组与文献中的分组对应起来。原题中的③-2(同时使用A试剂和NAA、乙烯)在原文献中未找到出处,眼尖的老师找到了一定留言要告诉我。
从实验4的结果可以看到,受体抑制剂的使用确实减轻了野生型的乙烯反应,这说明生长素与受体结合(或结合后开启的生长素信号通路)是触发乙烯反应的必要条件,此处对应原题第(3)问。围绕实验4的设问还有(2)-③,属于常规的实验对照分析,这里不做解读。
这里顺便吐槽一下(3)的设问方式:
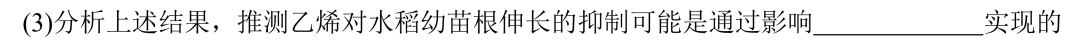
这一空的答案明明只看实验4就足够了,前半句却多了一句烟雾弹“分析上述结果”,这会让人误以为这一结论的得出需要综合考虑实验1-4,从而干扰解题。我第一次做这道题时,就因为多了这半句话,在这里就卡了很久去审视实验1-4之间的关系。
有老师可能会反驳,“分析上述结果”是因为实验2、3和4之间是并列关系,2和3的结果可以帮助排除一些其他机制的可能性(生长素合成/运输),最终取实验4的结论。这种说法只看题干好像说的通,但却解释不清楚一个问题:为什么实验2和3要引入m2突变体?在WT中去验证这些可能性不是更合理么?
要搞清楚这个问题,只看被删除了一些关键信息的题干是不够的,我们要去精读下原文献。参考原文写作中的表述,作者的逻辑应该是:既然实验1和4发现乙烯反应依赖于生长素信号途径,而实验室前期发现的一个水稻突变体m2又没有乙烯反应,这就有理由推测m2的突变基因影响了生长素的信号转导途径。所以,接下来的实验2和实验3的探究是围绕着解析m2突变基因的功能来展开的,然而原题干中却略去了这一点,在设问中把m2突变基因的功能验证放到了第一个科学问题底下,读起来略显怪异。

文献中提到了一些前人对拟南芥突变体的研究报道了在拟南芥中生长素的合成/运输基因突变会影响乙烯反应。由此,作者首先怀疑m2突变基因可能是参与了生长素的合成或运输等过程,进而影响生长素的信号转导,这才展开了实验2和3(读到这里读者会发现,即使植物生理背景出身的原文作者,在探究m2突变基因功能的时候也需要考虑多项前人研究。而19题题干中由于缺少对这些背景信息的交代,会导致不是相关背景出身的人在解题的时候会云里雾里,我凭什么知道要去比较生长素的合成量?凭什么要检测转运蛋白的表达量?)。
实验2的原假设m2的突变基因参与了生长素合成,实验3的原假设是m2的突变基因参与了生长素运输。
如果实验2的假设正确,那么m2突变体的IAA含量应该与野生型有差异(此处对应设问(2)-①):
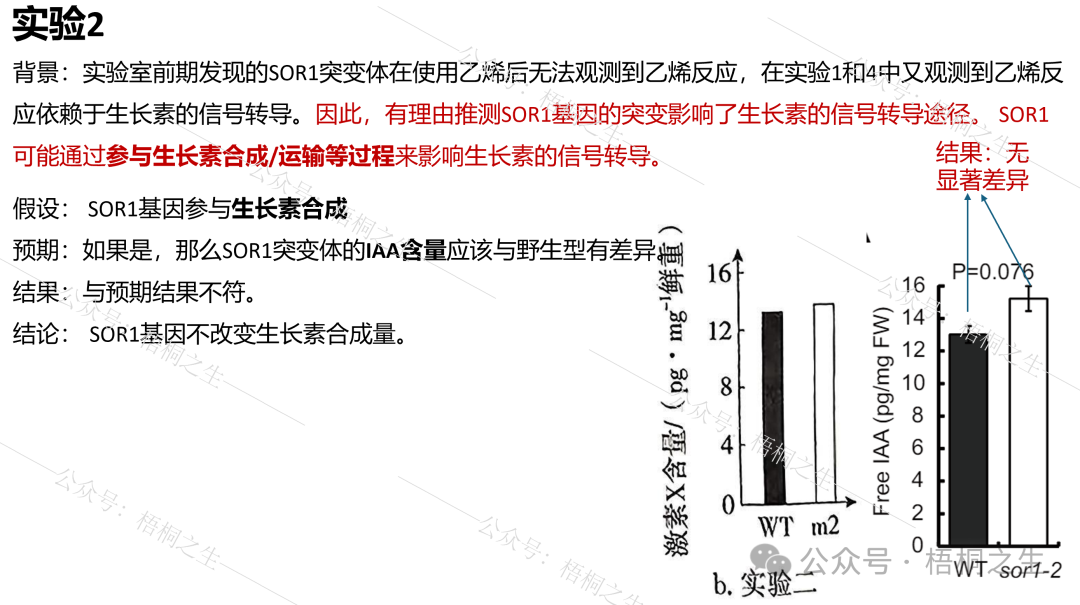
SOR1-2即题中的m2突变体
如果实验3的假设正确,那么突变体的生长素转运蛋白基因表达量应当与野生型有差异(此处对应设问(2)-②-2)。同时,对SOR1突变体使用可自由扩散的NAA(就可以不受转运蛋白的影响),应当能使突变体恢复乙烯反应(此处对应设问(2)-②-1):
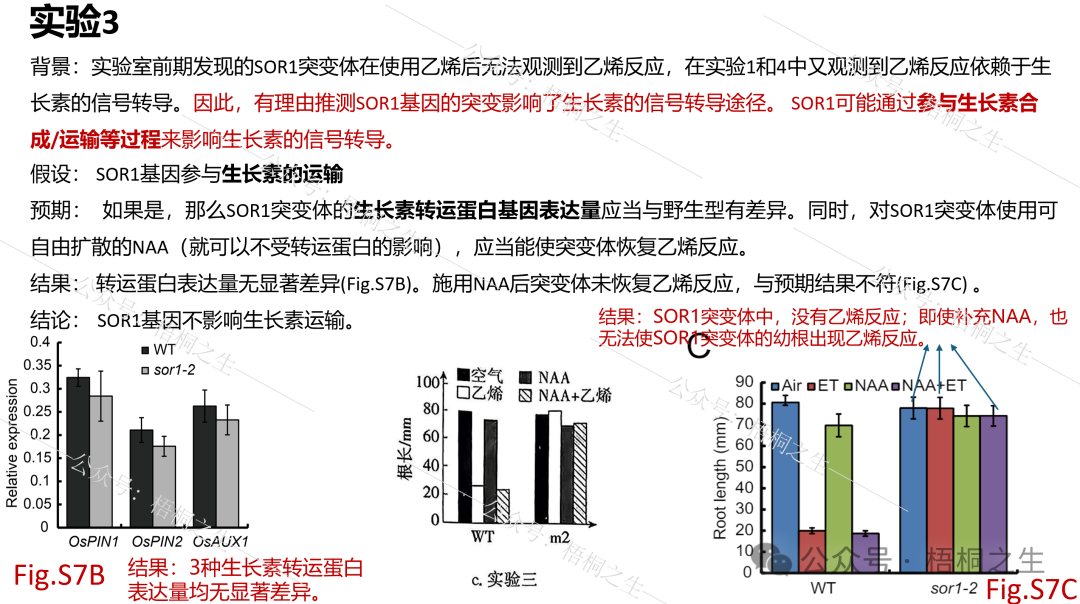
图中的OsPIN1、OsPIN2、OsAUX1即生长素转运蛋白。SOR1-2突变体即题中的m2突变体。
然而,从实验2和3的结果来看,都与预期不符。这就说明SOR1既不通过参与生长素合成、也不通过参与生长素运输来影响乙烯反应。至于SOR1的功能到底是什么,有兴趣的读者移步原文。
读到这里,细心的读者会发现实验1和4之间属于递进关系,两者都是为了探究生长素是如何参与乙烯反应的,因此实验1、4使用的都是野生型,与m2突变体没有任何关系: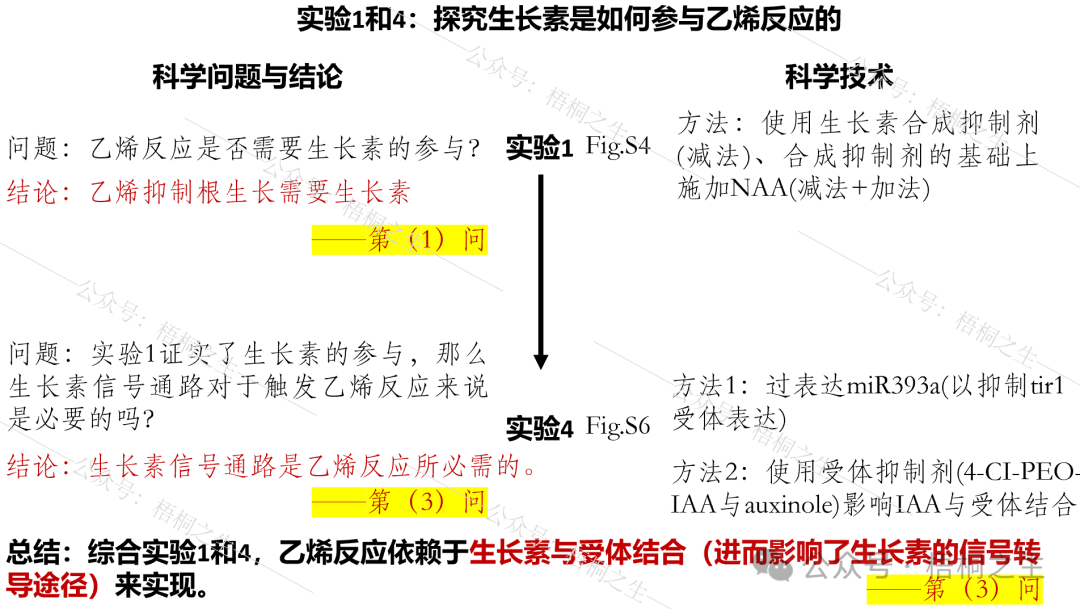
而实验2和3则是在前边2个实验结论成立的基础上,去进一步回答另一个科学问题,即m2突变基因的功能是什么,它如何影响生长素信号转导进而影响乙烯反应?2、3两个实验属于并列关系:

高考结束后,网上有一句吐槽“考完一科丢一科,考完生物丢本科”,要论哪道题的贡献最大,我猜19题应该当之无愧了吧?这道题都说难,到底难在哪呢,我结合自己做题时经历的困惑、以及读文献后反复琢磨再到逐渐明晰的过程,谈谈一点体会:
1.植物生理背景的缺失导致读懂题有一定困难。大家都知道生物行业隔行如隔山,即使都是生物学的博士,如果研究方向不同,两个人进行专业交流时都可能是鸡同鸭讲话。笔者并非植物生理背景出身,所阅读过的植物激素调节机制研究的相关文献也不多,这就导致要在短时间内把“乙烯反应、生长素以及m2突变体”以及“生长素合成、运输、信号转导等过程”等多个概念之间的关系理清楚,对于我一个老师来说尚且有困难,更别提考场上的学生了。我在想几十万考生中真的有人在解完19题后弄懂了这道题到底在干什么吗…如果没有,答案也是边写边猜的,那出这道题的目的是?
2.题干文字表述有瑕疵。在前边的文献梳理中提到,4个实验是围绕着2个相关的科学问题展开的,4个实验之间的关系既有递进关系,又有并列关系。这些信息如果笔者没有读原文献,只靠题干那4行文字恐怕根本看不出来。而原题是怎么处理这些信息的呢?把所有主要信息全部放在总题干中,2个科学问题只提及1个,紧跟题干的是四个摆在一起的实验,然后在设问中还有出现了一些诡异的、可能干扰分析的描述。这种情况下要人做出来真的有些强人所难,我猜这题的实际得分数据应该不怎么好看吧?



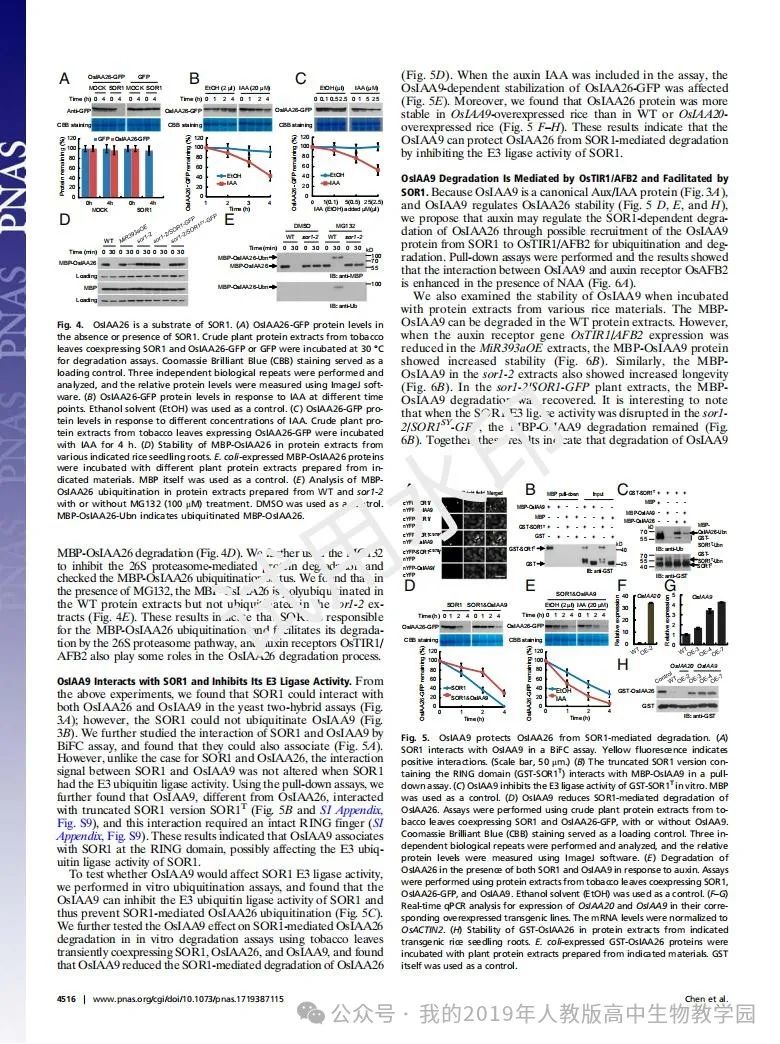







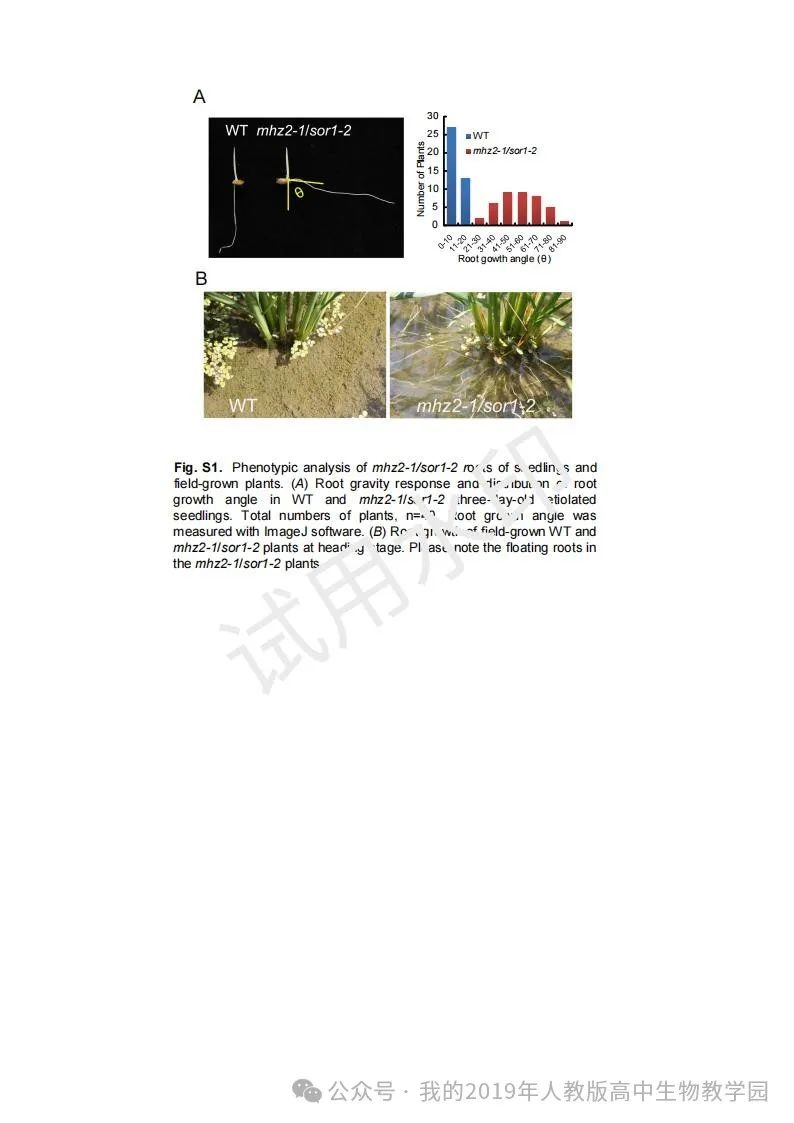


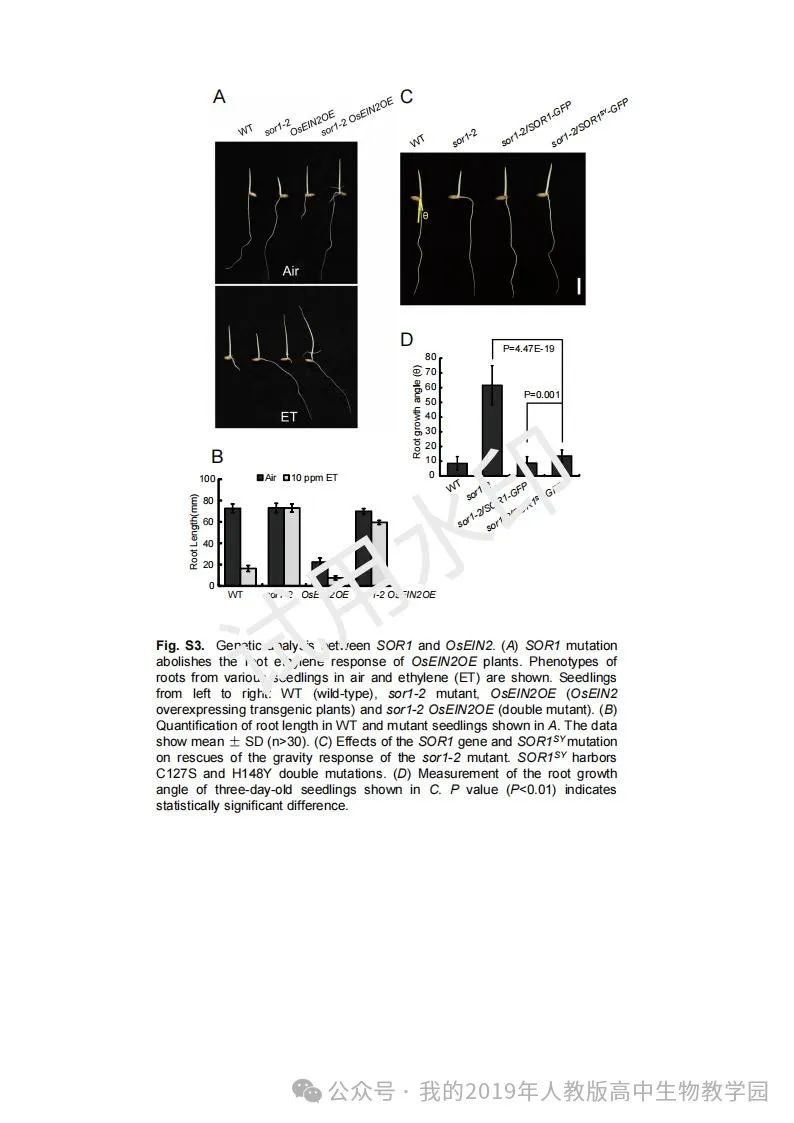
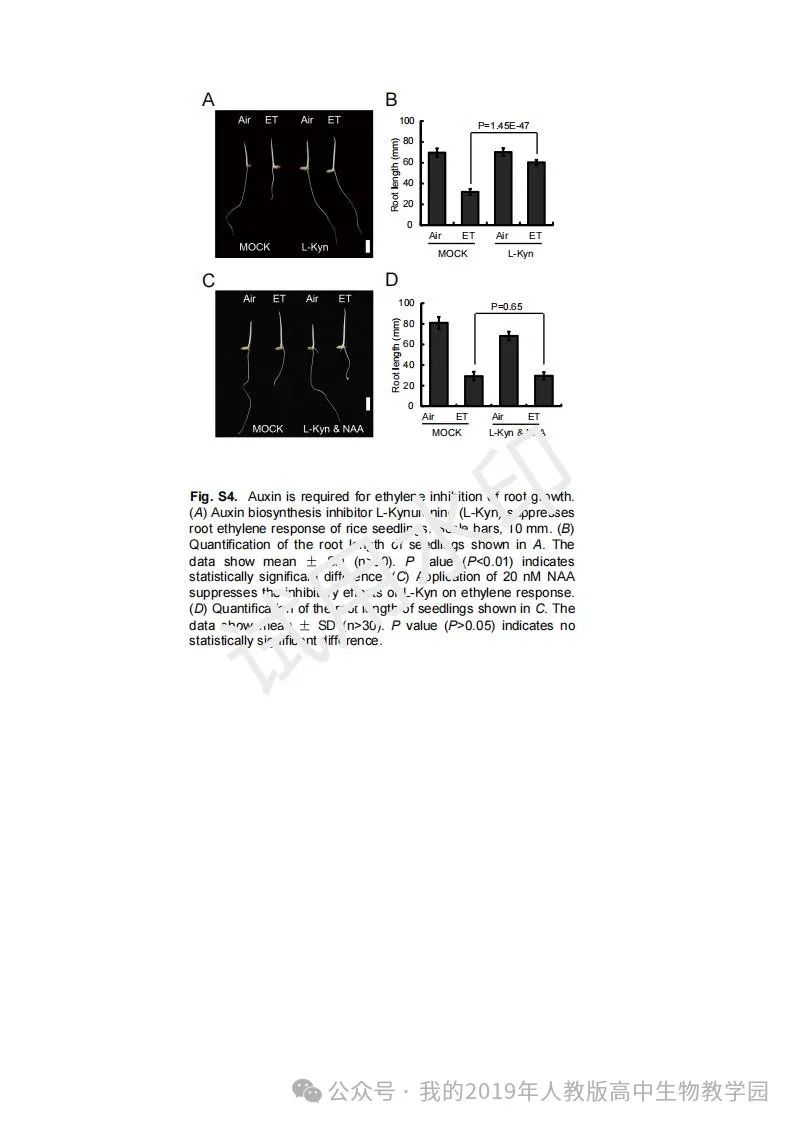
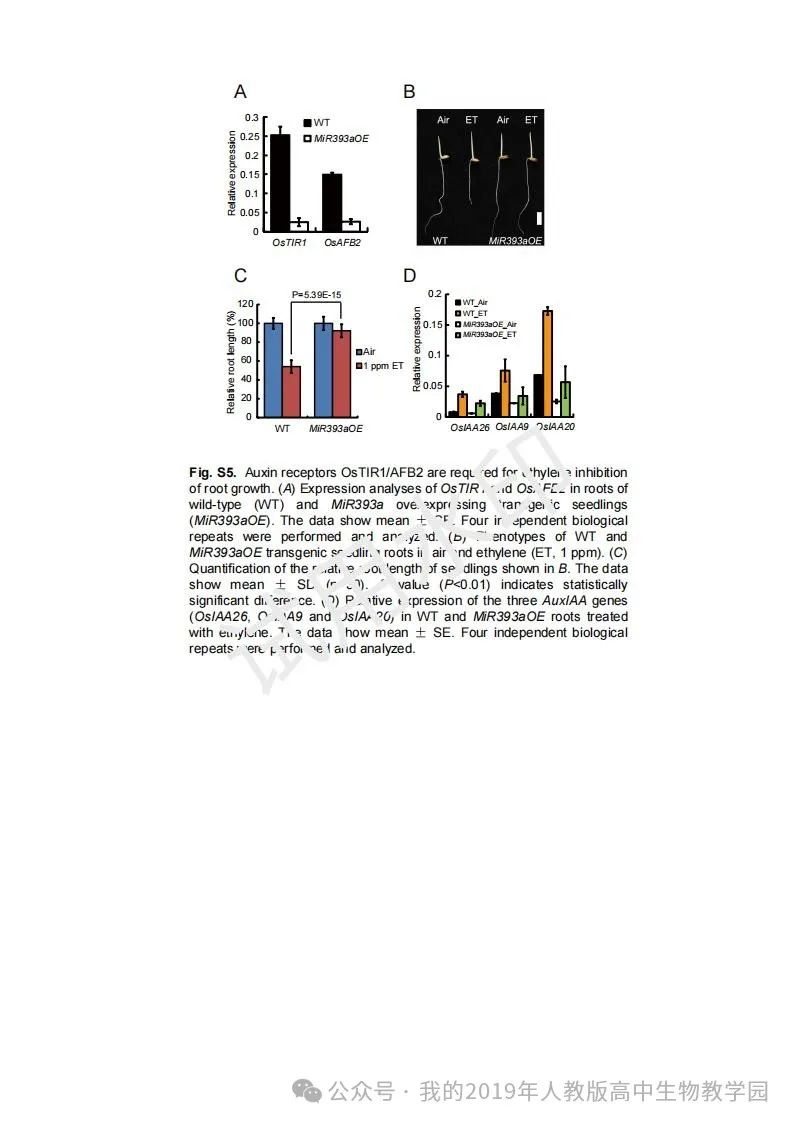
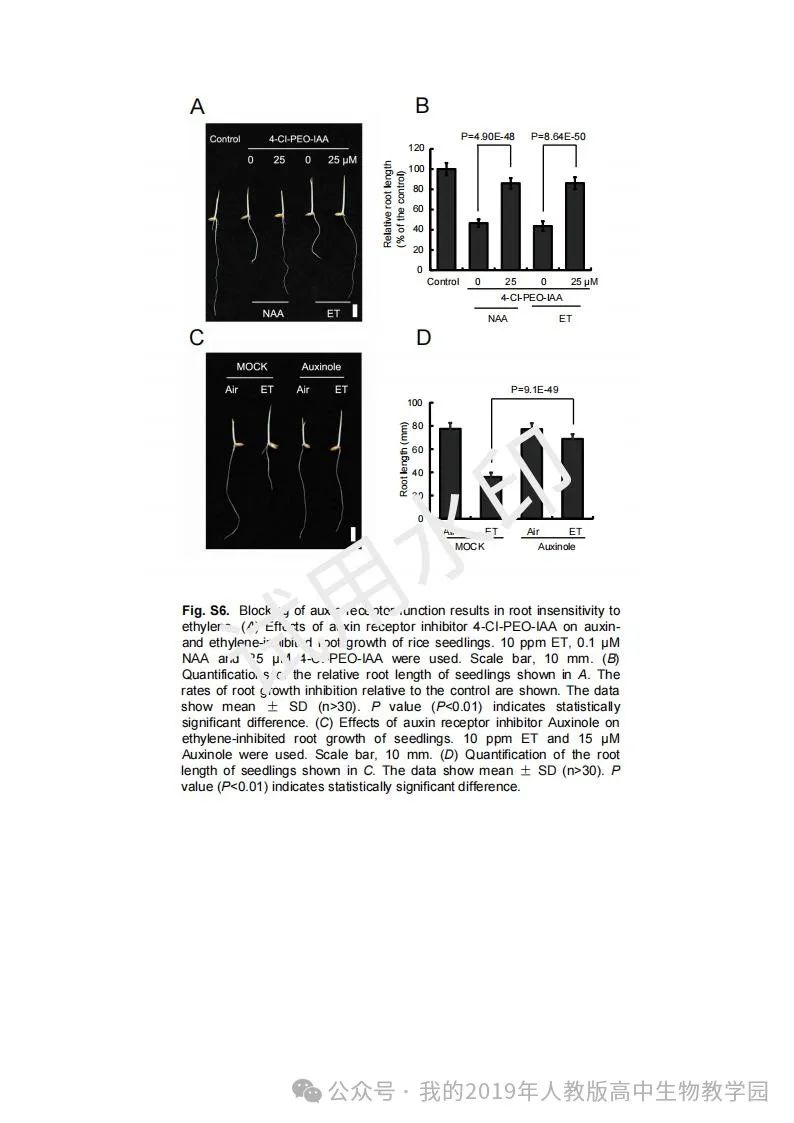
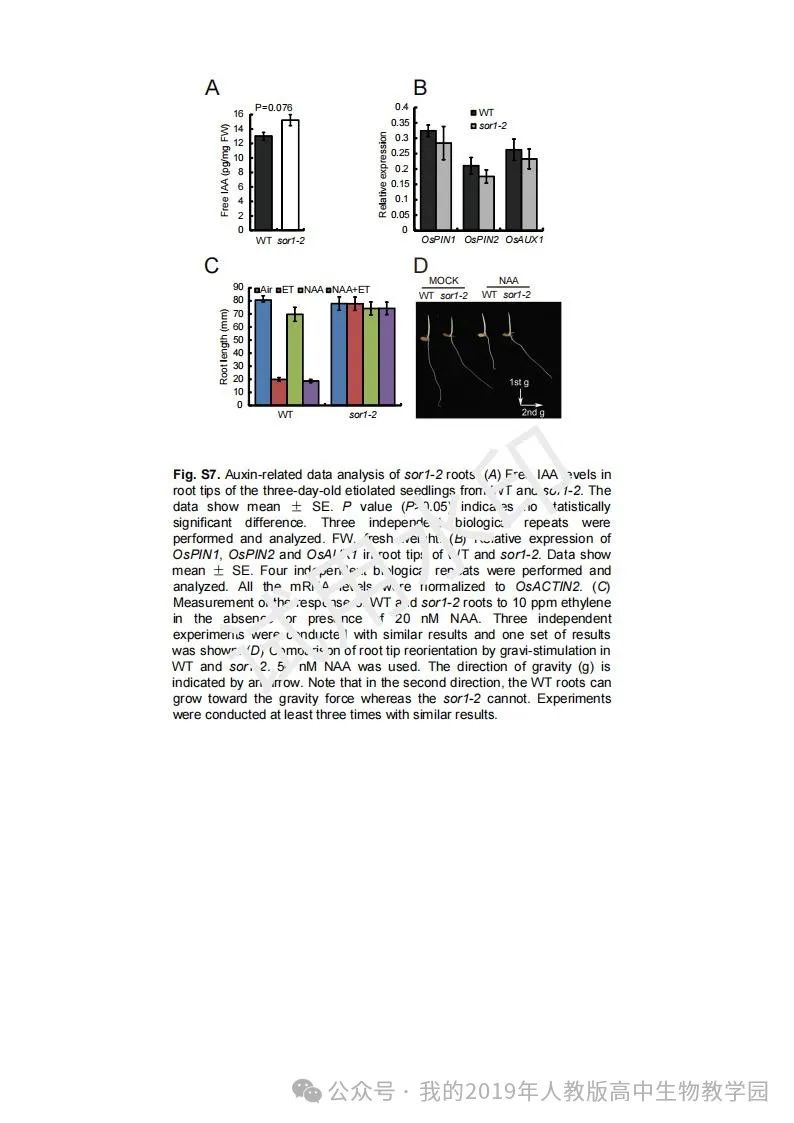
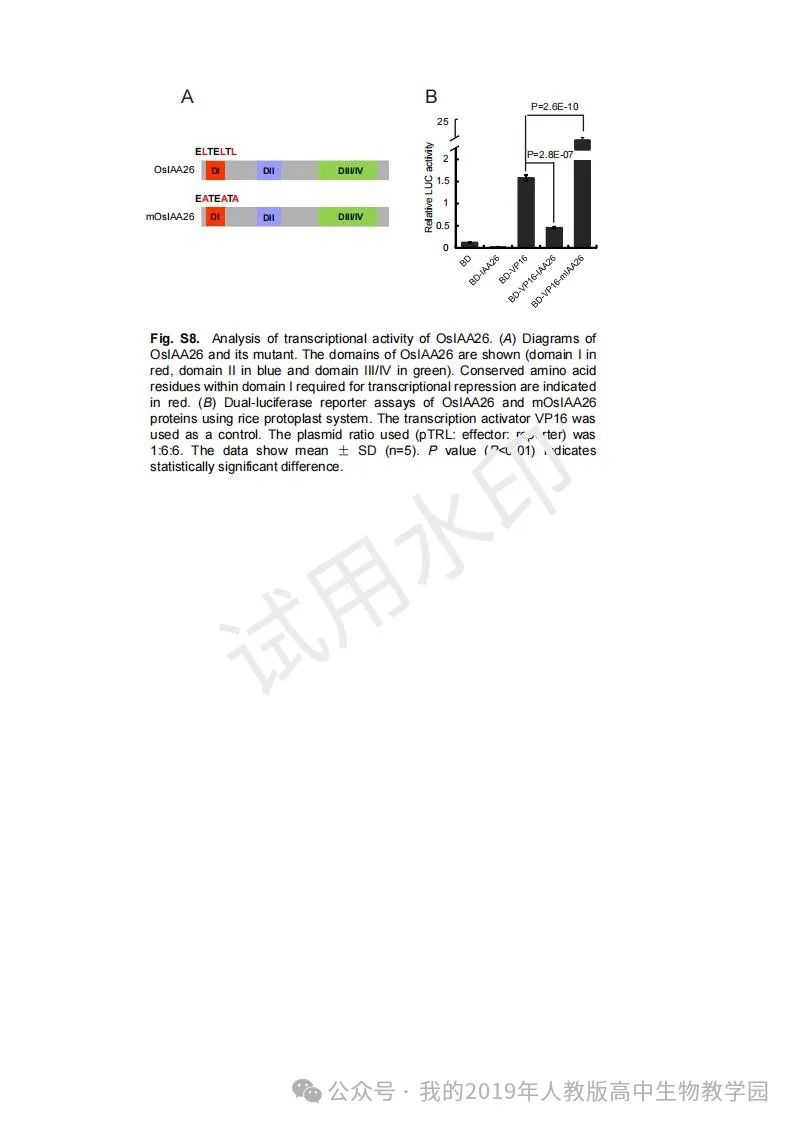
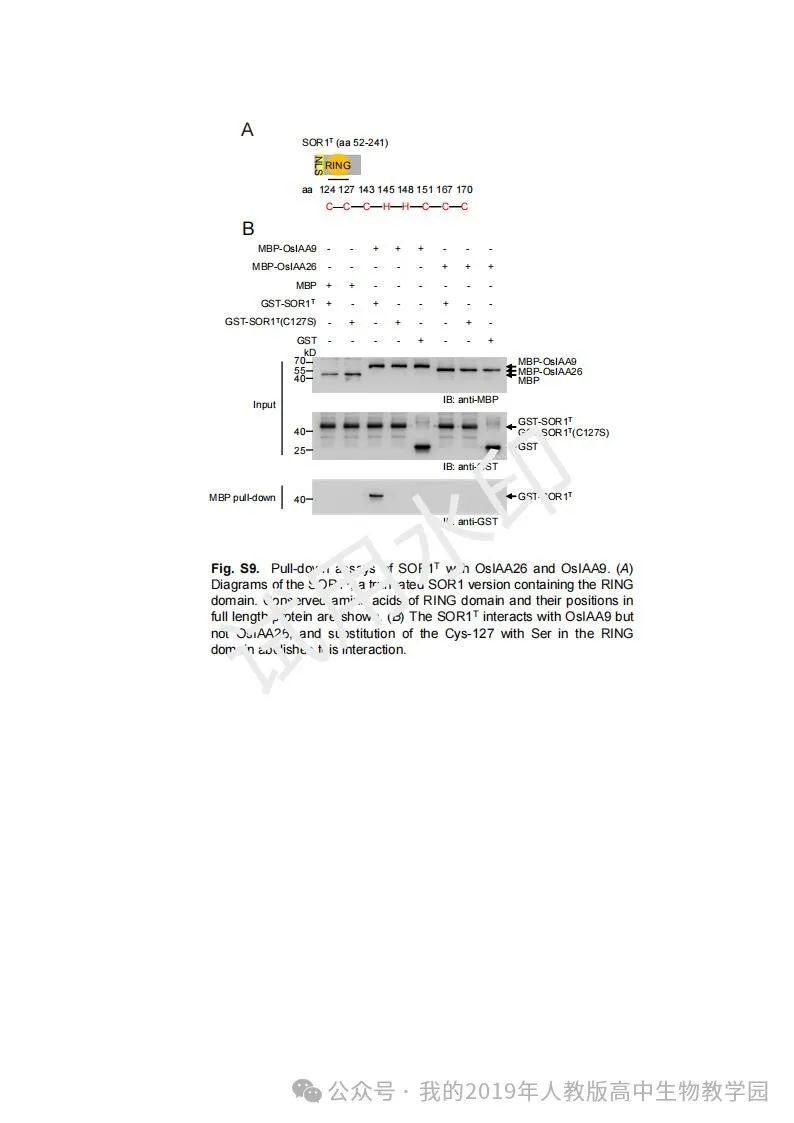
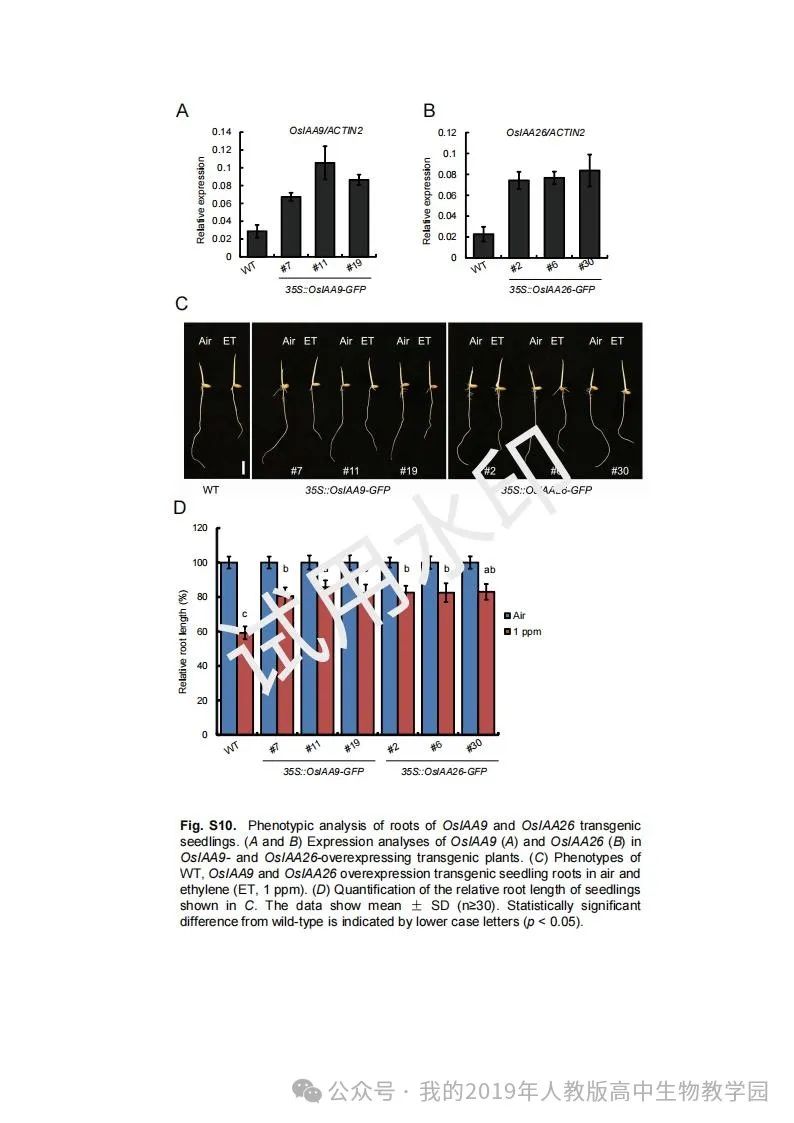
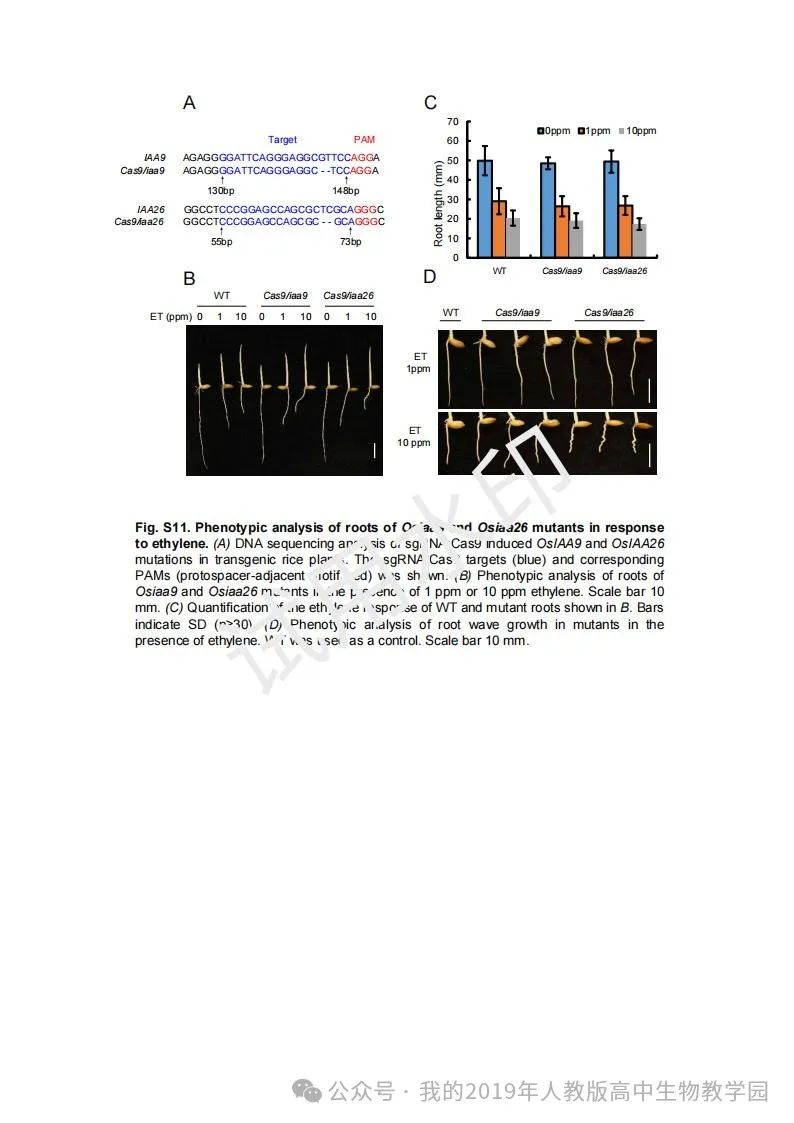



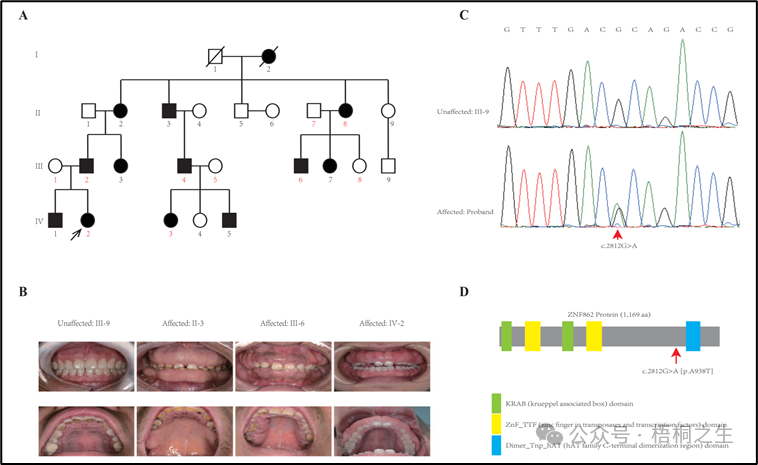
从给出的家系图来看,18题改编自22年发表在eLife的一篇文献:

第(1)问遗传方式+简单的概率计算,注意问对象是IV2(根据网络流传版本推测),没有任何关于婚配丈夫的信息。所以需要用数学上的全概率公式进行计算:

尽管这几年的高考已经弱化了计算,但并不代表生物学的研究中不能出现任何数理知识。况且这一问涉及的概率计算并不算复杂,即使课堂没有着重训练,数学课上应该也讲过。第(2)问则承接第(1)问表现型的基础上,继续深入探究致病基因及其机制,分子手段研究某个基因与表型的联系往往通过敲除/敲入来实现,今年深二模的遗传题曾有所涉及,因此本问不算特别新鲜。第(3)问与原文献研究内容无关,设问围绕应用层次,呼应了前几年南科大基因编辑婴儿事件,渗透社会责任感。
如果老师曾在课堂上提到过这一事件,第(3)问学生的答案至少在观点上不会出错。用学生的话说,上一个同意的人已经进去了:)。
总的来说这道人类遗传病的遗传题质量是不错的。设问环环相扣,遵循了人类遗传病的一般研究范式:从家系分析→发现致病基因和位点→机制推测。每一环节都有待探究的科学问题,也有特定的生物技术回答这些问题。
教学启示高三阶段,常听到要培养学生的科研思维,要让学生像科学家一样思考,因此课堂教学少不了科研情境。但情境的选择也有讲究,不能什么垃圾题目或素材都拿来用。这道题就是一个很好的素材,课堂上完全可以参考原文献把情境铺开(有兴趣的老师推荐去读读原文,下图是笔者梳理的原文献研究过程):
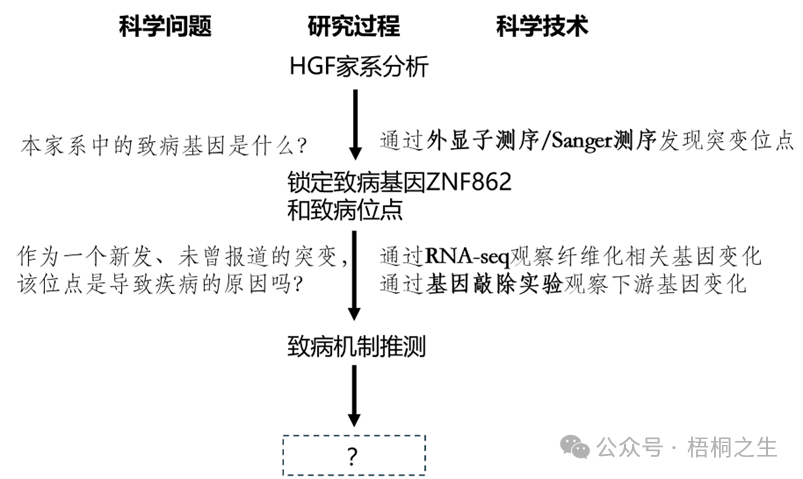
可以看出命题人有意省略了一些实验技术和结果,是降低了难度的。
对于这类比较有代表性的科研情境,学生解题的同时,也对这类情境的研究范式有初步了解。久而久之,也就掌握了研究这类问题的一般方法,这不就是科研思维么。这里抛砖引玉在原试题的基础上多设几个问题,供读者思考:
1.你认为这项研究的价值是什么?上图中的虚线方框该填什么?
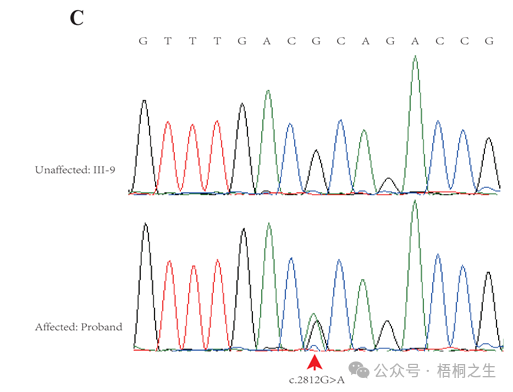
2.根据下图测序结果,推测致病位点及突变的类型是?
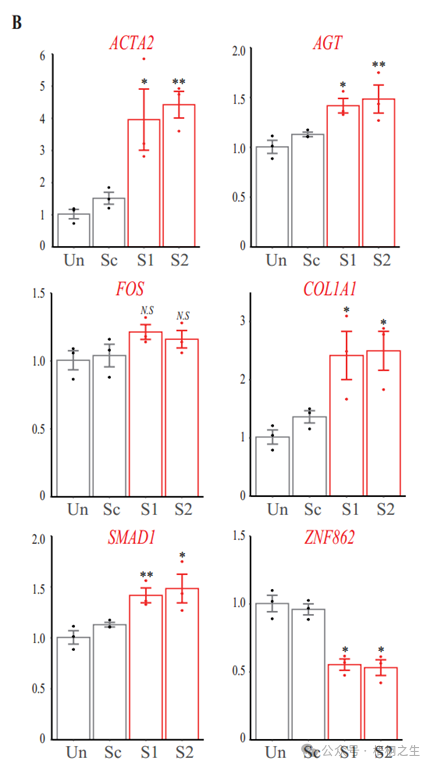
3.敲低ZNF862后,ACTA2、FOS、SMAD1、AGT和COL1A1等促纤维化基因表达量变化如下图所示,据此推测ZNF862影响疾病的机制可能是?
原始文献

一个新的基因ZNF862导致遗传性牙龈纤维瘤病
摘要
遗传性牙龈纤维瘤病(HGF)是最常见的牙龈纤维瘤病的遗传形式,其特征是牙龈局部或全身性过度生长。目前已确定两个基因(SOS1和REST)以及四个位点(2p22.1、2p23.3-p22.3、5q13-q22和11p15)以显性遗传模式与HGF相关。在这里,我们报告了来自一个四代中国家庭的13个常染色体显性HGF个体。全外显子组测序和进一步的遗传共分离分析对三代家庭成员进行。在锌指蛋白862基因(ZNF862)中发现了一个新的杂合错义突变(c.2812G > A),该突变在种群中未见。功能研究支持ZNF862的生物学作用,增加促纤维化因子,特别是COL1A1的合成,从而导致HGF。在这里,我们首次确定了ZNF862与HGF相关的生理作用。
引言
牙龈纤维瘤病(GF)是一种罕见的疾病,其特征是良性的,非出血性的,局部的或全身性的纤维扩大,游离和附着的牙龈进展缓慢。GF可能并发癫痫、多毛症和智力低下(Balaji and Balaji, 2017;Snyder, 1965),也可以发展为综合征的一部分,如Cowden综合征(Witkop, 1971)、Zimmerman-Laband综合征(Guglielmi等人,2019)、Cross综合征(Poulopoulos等人,2011)、Rutherford综合征(Häkkinen和cisszar, 2007)、Ramon综合征(Suhanya等人,2010)、Jones综合征(Gita等人,2014)、Costello综合征(Hennekam, 2003)、电性amelia综合征(Morey和Higgins, 1990)和透明纤维瘤病综合征(Hamada等人,1980);它也可能是由与口腔卫生有关的牙龈炎症、青春期、怀孕或钙通道阻滞剂(硝苯地平和维拉帕米)等常见药物的副作用引起的(Livada和Shiloah, 2012);通常表现为非综合征性遗传性牙龈纤维瘤病(HGF) (Jorgenson and Cocker, 1974)。HGF是GF最常见的遗传形式,通常以常染色体显性遗传模式传播。然而,以前也有零星病例和常染色体隐性遗传谱系的描述(Majumder等人,2013年)。
如前所述,迄今为止,已鉴定出四个位点(2p22.1 [MIM: 135,300],5q13-q22 [MIM: 605,544],2p23.3-p22.3 [MIM: 609,955]和11p15 [MIM: 611,010])与HGF相关(Xiao等,2001;Ye et al .,2005;Zhu等,2007;Xiao et al .,2000);此外,在一个巴西家庭中,SOS1的杂合移码突变(MIM: 182530)已被确定为常染色体显性HGF的病因(Hart等,2002);REST中的蛋白质截断突变(MIM: 600571)已被报道为三个独立家族中常染色体显性遗传模式HGF的遗传原因(Bayram等,2017)。HGF的估计发病率为每17.5万人中有1人(Ahmed and Ali, 2015),男女发病率相等(Gawron et al, 2016;Odessey et al.,2006)。由于美观和功能方面的考虑,牙周手术包括牙龈切除术、牙龈成形术和皮瓣手术可以应用于HGF患者,然而由于潜在的遗传易感性,增生的复发率相对较高(Chaurasia, 2014;Zhou et al .,2011)。因此,探索HGF的遗传原因和病因学,对精确的遗传诊断和潜在的基因治疗发展有重要意义,以帮助那些希望避免痛苦和改善生活的个体。我们报告了一个中国大家庭,有13名临床诊断为非综合征性HGF的患者,以及12名未受影响的家庭成员。对这个大家族的遗传分析导致了一个新基因ZNF862的杂合突变的鉴定。
结果与讨论
图1A显示了这个带有HGF的四代家系。目视检查显示所有受影响的成员有严重或轻度的广泛性GF,而没有任何未受影响的成员,来自这个家庭。根据我们的调查,这个家庭中没有人接触过可能导致GF的药物。所有家庭成员均未发现其他先天性异常。该家族所有个体的临床资料汇总于表1,图1B为每代1名对照和3名患者的临床照片。从临床角度看,先证者(IV-2)在牙周手术包括牙龈切除、牙龈成形术、皮瓣手术后第二年出现轻度增生复发;而患者II-2有额外的临床表现(高血压,未接受钙通道阻滞剂治疗)。
根据先前描述的方案(Zhang et al .,2015),对先证和该家族的其他9名成员(图1A中以红色表示的数字)进行全外显子组测序(WES)。在分析了启动子区、外显子、剪接位点、内含子和非翻译区(UTR)等基因的所有单核苷酸变异(snv)和indel后,在10名接受WES治疗的家庭成员中,仅鉴定出3个变异ATP7B (c.3403G > A)、CDADC1 (c.83-13G > T)、ZNF862 (c.2812G > A)与表型共分离。为了进一步筛选和验证该家系中与表型共分离的潜在致病变异,采用常规PCR进行sanger测序评估。最终,只有ZNF862 (c.2812G > A)被证实与该家族所有23名成员的表型共分离,没有任何其他变异,如图1C所示。
ZNF862基因含有8个外显子,编码1169个氨基酸的蛋白,位于染色体7q36.1上。我们注意到ZNF862没有映射到先前报道的与HGF相关的任何一个位点。ZNF862是一种预测的细胞内蛋白,据我们所知,其功能尚未明确,作为锌指蛋白,它可能参与转录调节。ZNF862在组织中普遍表达(Fagerberg et al .,2014),在不同的生理条件下可能发挥不同的作用。Schwartz等人,2019提出了ZNF862在基质生成的化生癌中可能的作用。Peng等人,2018年报道了ZNF862与儿童ige介导的i型超敏反应相关。在我们的研究中,wes鉴定的杂合变异体(p.A938T)位于ZNF862蛋白Dimer_Tnp_hAT (hAT家族c端二聚化区)结构域的上游附近(图1)。
HGF的病理生理机制在很大程度上仍然难以捉摸。然而,细胞外基质的过量产生,特别是主要成分胶原I型(COL1A1),可能是牙龈成纤维细胞过度生长表型的原因;同时,TIMP-1合成的增加似乎与HGF牙龈成纤维细胞中COL1A1的过度积累有关(Gawron等,2018;Roman-Malo et al, 2019)。此外,Martelli-Junior等报道TGF-β1和IL-6在牙龈成纤维细胞中过表达,在增加COL1A1及其他特异性生长因子的合成中起关键作用(Martelli-Junior等,2003)。Han等(2019)对HGF患者和对照组的转录组学分析系统地表明,TGF-β/SMAD信号通路与颅面发育过程的调控网络连接有助于临床病理表现的分子机制。
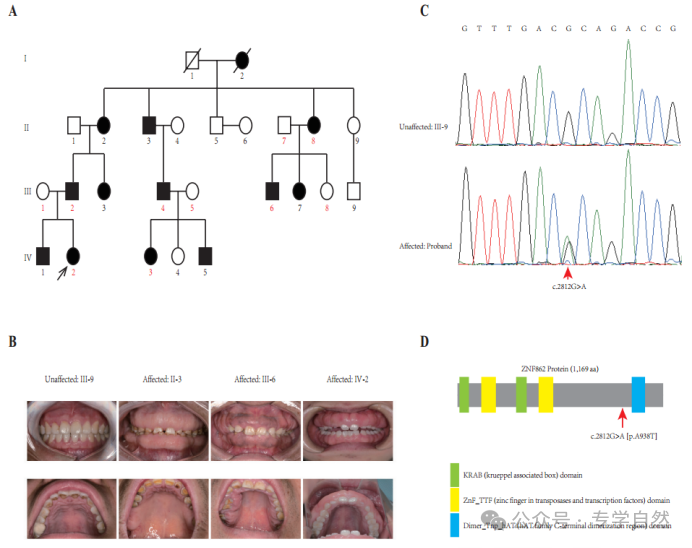
图1所示。谱系和共分离分析。(A)黑色箭头表示先证者。在这个家族中,受影响的个体用黑色填充框表示。先证者和其他9个成员(数字用红色表示)进行全外显子组测序(WES)。(B)本家族受影响成员与未受影响成员的口腔内检查显示牙龈过度生长的照片。(C)在这个家族中,所有受影响的个体都携带杂合变异体(C . 2812g > A),而未受影响的个体为野生型。(D) ZNF862蛋白推定结构域的结构示意图和新变异的定位(红色箭头)。绿色矩形表示KRAB (krueppelassociated box)域;黄色矩形表示ZnF_TTF(转座酶和转录因子锌指)区域;蓝色矩形表示Dimer_Tnp_hAT (hAT家族c端二聚化区)域。
为了探索与ZNF862突变相关的潜在机制,根据先前描述的方案进行了RNA测序(Liang et al .,2019)。原代成纤维细胞是从两名接受牙龈切除术的患者(IV-1和IV-2)的纤维化牙龈标本中分离出来的,使用标准外植体培养,如先前所述(de Andrade等人,2001年),而对照细胞来自四个独立的年龄和性别匹配的对照组,他们接受了修复目的的冠延长手术。补充文件1总结了患者的基因表达谱和对照组的变化,显示了每个基因的样品平均每百万映射读取数(CPM),表达折叠变化(FC)及其统计显著性错误发现率(FDR)。我们对可能参与HGF的已证实的纤维化基因(Gao et al, 2019)的表达谱进行了研究(图2A)。与对照组相比,这些基因的一部分,包括COL1A1、TGFB1/2和SMAD1,在HGF成纤维细胞中表达上调;而另一部分下调,提示我们研究中的特殊HGF归因于相应的促纤维化因子,包括TGF-β/SMAD1信号通路和COL1A1。除了RNA测序外,对照1体外培养的牙龈成纤维细胞进行了腺病毒介导的靶向ZNF862的短发夹RNA (shRNA)递送。研究了四个最重要的基因ACTA2、FOS、SMAD1、AGT和COL1A1在shRNA处理下的表达情况(图2B),对照组牙龈成纤维细胞中ZNF862的敲低导致原纤维化基因上调,与患者的表达情况相似。本研究推测,患者的ZNF862突变与ZNF862 shRNA相似,削弱了该蛋白的功能,促进了特殊的促纤维化基因的表达,从而导致HGF性状的产生。同时,我们也注意到致病基因SOS1出现了轻度下调,这可能与表型有关(图2和图2 -图补充3)。然而,在对照牙龈成纤维细胞中,ZNF862的下调在培养5天的时间内并没有显著刺激其增殖(图2 -图补充2),这可能是由于时间太短,无法观察到复杂的生理效应。

为了筛选患者和对照组之间的所有差异表达基因(DE),我们量化了表达变化超过两倍的基因(|log2 FC|≥1.0),由于整个样本数量少(N = 6),我们采用FDR < 0.05作为截止值。最后,HGF患者和对照组的转录组比较得到597个DE转录本,其中355个表达上调,242个表达下调(图S1)。其中,DE上调最多的100个基因和DE下调最多的100个基因(Supplementary file 2)如图2C所示,并对前10个基因进行了图示。这些DE基因可能与HGF性状相关。显示了所有DE基因中20条最显著富集的功能通路(图2D)。其中,硫辛酸代谢最为富集;值得注意的是,MAPK信号通路与IL-6通路有关,此外,图中几个感染相关的簇与IL-6诱导的自噬通路有关。
综上所述,我们推测ZNF862可能作为转录抑制因子发挥作用,因为ZNF862具有锌指dna结合结构域,同时这种错义突变可能减弱ZNF862的生物学功能,从而增强TGF-β/SMAD1信号通路,从而增加促纤维化因子,特别是COL1A1在牙龈组织中的积累,导致HGF的发生。
然而,这种新突变导致HGF的潜在机制仍然是一个谜,没有更具体的实验设计来探索ZNF862的生物学功能。进一步的研究有望扩大ZNF862的突变谱及其与表型的关联,探讨ZNF862在HGF过程中的生理作用。
在这项研究中,我们在一个中国四代家庭中发现了一个导致常染色体显性HGF的新基因ZNF862的错义变异(c.2812G > a)。功能研究支持ZNF862的生物学作用,增加促纤维化因子,特别是牙龈中COL1A1的合成,从而导致HGF。ExAC (Exome Aggregation Consortium)数据库、1000 Genomes和Genome Aggregation database的报告显示,该变异在大人群中是不存在的。赔率(LOD)分数的对数根据优势隔离计算为3.6。根据美国医学遗传学和基因组学学院(ACMG)和临床基因组资源(ClinGen)的联合标准制定指南(Oza等,2018;Richards等人,2015),错义变异被评为“致病性”,证据规则如下:PP1_Strong (LOD > 3),PS3(体外功能研究支持)和PM2(对照组中不存在)。我们可以推测,这一发现可能会为精确的基因诊断和潜在的治疗发展提供线索,帮助人们更好地生活。
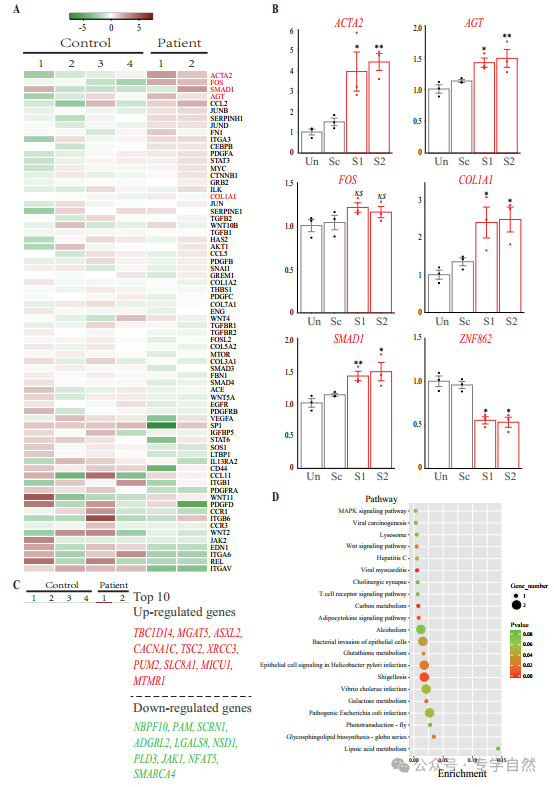
图2。RNA测序和敲低分析。(A) RNA测序中70个已证实的纤维化基因表达谱的热图。行代表基因,列代表样本。热图是基于平均归一化的颜色编码;红色表示高表达值,绿色表示低表达值。(B)实时PCR分析ACTA2、FOS、SMAD1、AGT、COL1A1和ZNF862 mRNA丰度在未处理(Un)、腺病毒传递的乱码器(Sc)、腺病毒传递的ZNF862短发夹RNA (shRNA) 1 (S1)和腺病毒传递的ZNF862中。shRNA 2 (S2)组。结果以平均值±SEM给出,圆点表示基因表达水平的相对值。*p < 0.05,**p < 0.01与Sc组比较。N.S.的意思是不重要。(C) RNA测序中100个差异表达基因上调最多和100个差异表达基因下调最多的热图。色键与(A)相同。(D) DE基因功能注释图,绘制了DE基因功能最富集的20个位点。功能通路富集试验基于KEGG数据库。富集表示DE基因在指示通路中的比例。
材料与方法样品采集与WES
该家庭的参与者来自南京市口腔医院。从外周血中提取基因组DNA。对该家族的先证者和其他9名成员(图1A中以红色表示的数字)进行WES检查。每个样本目标区域的平均WES测序深度大于100,每个样本的目标碱基覆盖率达到95%以上的20倍。测序读数通过内部程序与人类基因组参考序列(hg19)对齐。使用Genome Analysis Toolkit (GATK)对snv和indel进行调用;采用遗传变异标注(ANNOVAR)进行标注。一个自定义Perl脚本(源代码1)用于检索与这个家族的继承模型相对应的变体,这些变体不会出现在未受影响的个体中。所有的WES数据都经历了相同的质量控制过滤和修剪程序,以最大化奇偶性。
转录组测序
原代成纤维细胞是从接受牙龈切除术的患者(IV-1作为患者1和IV-2作为患者2)的纤维化牙龈标本中分离出来的,使用标准外植体培养,而对照细胞来自年龄和性别匹配的独立对照组(分别为25岁和26岁的两名男性作为对照1和2,21岁和23岁的两名女性作为对照3和4)。分别)为修复目的接受冠延长手术的患者。
使用Trizol从培养的成纤维细胞中提取总RNA样本,使用Bioanalyzer 2100系统(Agilent Technologies, CA)的RNA Nano 6000 Assay Kit评估RNA完整性。以每个样品1μg的RNA为输入物制备RNA样品。简单地说,利用聚t寡聚磁珠从总RNA中纯化mRNA。在第一链合成反应缓冲液中,用二价阳离子在高温下进行裂解。利用随机六聚体引物和M-MuLV逆转录酶(RNase H-)合成首链cDNA。随后使用DNA聚合酶I和RNase h进行第二链cDNA合成,剩余的悬垂部分通过核酸外切酶/聚合酶活性转化为钝端。DNA片段3′端腺苷化后,连接具有发夹环结构的接头,准备杂交。为了优选长度为370-420 bp的cDNA片段,使用AMPure XP系统(Beckman Coulter, Beverly, MA)对文库片段进行纯化。然后用Phusion高保真DNA聚合酶、通用PCR引物和Index引物进行PCR。最后,用AMPure XP系统纯化PCR产物,并在Agilent Bioanalyzer 2100系统上评估文库质量。文库制备在Illumina Novaseq平台上测序,产生150 bp的成对末端reads。
原始数据通过内部生物信息学工作流程进行处理。使用Hisat2 v2.0.5将配对的末端clean reads与参考基因组ensembl release-97 (http://asia.ensembl.org/Homo_sapiens/ Info/Index)比对,使用featurecots v1.5.0-p3对每个基因的reads数进行计数。然后根据每个基因的长度和该基因所映射的reads数计算每个基因的CPM和每千碱基转录本每百万映射reads (FPKM)的片段数。每个样本至少产生了4500万个干净的映射读数。各样品经过滤后的清洁碱基均大于6.7千兆碱基,有效率大于99.7%,Q20碱基对各样品的有效率大于94%。每个样品的GC含量范围为51% ~ 55%。采用edgeR R软件包(3.22.5)对两组(患者和对照组)进行DE分析。使用Benjamini和Hochberg的方法来调整所得的p值以控制FDR。校正后的p值为0.05,绝对FC值为2作为显著DE的阈值,我们使用clusterProfiler R软件包对KEGG通路中DE基因的统计学富集进行检验,校正后的p值小于0.05为显著富集。KEGG是一个用于理解生物系统的高级功能和实用程序的数据库资源(http://www.genome.jp/kegg/)。
论文下载链接:https://pan.baidu.com/s/1VzArZeeRv6iiffBpvK40yA?pwd=kcms 提取码:kcms

本文由小编发布,不代表一本线高考网立场,转载联系作者并注明出处:https://www.yibenxian.com/tiku/57366.html
